Artículo Original
Prospección de los coccinélidos asociados al cultivo de maíz (Zea mays L.)
Prospecting of coccinelides associated with maize cultivation (Zea mays L.)
 Nexar Emiliano Vega Lucas nexarvega24@hotmail.com
Ariolfo Leonardo Solís Bowen ariolfo.solis@utm.edu.ec
Dorys Terezinha Chirinos Torres dchirinos@utm.edu.ec
Diego Rodolfo Perla Gutiérrez diego_perla_g@yahoo.com.pe
Alex Gabriel Delgado Párraga alex.delgado7521@yahoo.com
Sofía Lorena Peñaherrera Villafuerte loren80@gmail.com
Nexar Emiliano Vega Lucas nexarvega24@hotmail.com
Ariolfo Leonardo Solís Bowen ariolfo.solis@utm.edu.ec
Dorys Terezinha Chirinos Torres dchirinos@utm.edu.ec
Diego Rodolfo Perla Gutiérrez diego_perla_g@yahoo.com.pe
Alex Gabriel Delgado Párraga alex.delgado7521@yahoo.com
Sofía Lorena Peñaherrera Villafuerte loren80@gmail.com
Prospección de los coccinélidos asociados al cultivo de maíz (Zea mays L.)
Siembra, vol. 11, núm. 1, e6021, 2024
Universidad Central del Ecuador

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Recepción: 10 Enero 2024
Revisado: 21 Febrero 2024
Aprobación: 29 Febrero 2024
Resumen: El maíz es considerado el tercer cultivo de grano más importante del mundo debido a su interés económico, alimentario e industrial. A lo largo de su fenología este cultivo es atacado por plagas y enfermedades, que pueden generar efectos adversos sobre la producción. Para el control de estos problemas, generalmente se acude al manejo convencional, los cuales ocasionan desequilibrios ecológicos. Por esta razón, es necesario considerar alternativas sostenibles fundamentadas en el conocimiento de la entomofauna benéfica. Esta investigación tuvo por objetivo realizar un análisis prospectivo de los coccinélidos asociados al cultivo de maíz en el cantón Santa Ana, Ecuador. Para su efecto, se evaluaron los coccinélidos presentes en dos condiciones distintas: con y sin aplicación de insecticida. Se utilizaron cinco técnicas de muestreo, de acuerdo con el protocolo de investigación, y se procedió con la clasificación taxonómica. Los datos fueron interpretados mediante estadística descriptiva; además, se valoró la diversidad alfa y beta. En el estudio se encontraron 297 individuos, congregados a: 1 orden,1 familia, 13 géneros y 11 especies. La mayor abundancia se presentó en condiciones no intervenidas, determinándose que la técnica de trampeo más eficiente para su captura fueron las trampas cromáticas. Este tipo de investigación es importante para derivar programas de control biológico en cultivos de importancia económica.
Palabras clave: control, convencional, diversidad, entomofauna, trampas.
Abstract: Maize is considered the third most important grain crop in the world due to its economic, food and industrial interest. Throughout its phenology this crop is attacked by pests and diseases, which could have adverse effects on production. In order to control these problems, conventional management is generally used, causing ecological imbalances. For this reason, it is necessary to consider sustainable alternatives based on the knowledge of beneficial entomofauna. This research aimed to conduct a prospective analysis of the coccinelides associated with maize cultivation in the canton of Santa Ana, Ecuador. For this purpose, the coccinelides present under two different conditions were evaluated; with and without insecticide application. Five sampling techniques were used according to the research protocol and taxonomic classification was carried out. The data were interpreted using descriptive statistics, in addition, alpha and beta diversity was assessed. In the study, 297 individuals were found, gathered to: 1 order,1 family, 13 genera and 11 species. The greatest abundance was found in undisturbed conditions, determining that the most efficient trapping technique for their capture was the use of chromatic traps. This type of research is important to derive biological control programs in economically important crops.
Keywords: control, conventional, diversity, entomofauna, traps.
1. Introducción
El maíz (Zea mays L.) representa el primer cultivo de grano más importante en el mundo, debido a sus aportes en varios contextos: alimentario, económico, social y ambiental, es decir, forma parte de la dieta humana y animal, genera ingresos económicos, constituye la base para la elaboración de subproductos a nivel industrial, crea plazas de trabajos para muchas familias que dependen de este rubro mejorando su calidad de vida y también contribuye a la producción de biocombustibles (Tanumihardjo et al., 2020; Erenstein et al., 2022).
De acuerdo con la base de datos de la FAO se estimó que la producción mundial de maíz en el año 2021 fue de 1.210.235.135 tn, destacándose Estados Unidos y China como los principales países productores (FAOSTAT, 2023). En Ecuador, la superficie cosechada fue de 366.138 ha con una producción obtenida de 1.699.369 tn, sobresaliendo Los Ríos, Manabí y Guayas como las principales provincias productoras a nivel nacional (Instituto Nacional de Estadística y Censos [INEC], 2023).
A lo largo de su ciclo esta gramínea puede ser afectada por plagas y enfermedades que dañan diferentes órganos de la planta, las cuales podrían ocasionar pérdidas del rendimiento cercanos al 30 %; en este sentido, se han reportado más de 70 especies de insectos plagas asociados al maíz; sin embargo, seis especies tienen importancia económica (Hernández-Trejo et al., 2019). Para disminuir su incidencia se utilizan principalmente insecticidas químicos (Barreto Carbajal et al., 2022), sin considerar que su uso indiscriminado puede generar daños a la salud de los agricultores y consumidores, los cuales han sido documentados en Ecuador (Fernández Aravena, 2021; Vargas Sunta y Moyano Calero, 2022). Adicionalmente causan desequilibrios ecológicos de algunas plagas agrícolas derivándose vertiginosos incrementos poblacionales, problemas de resistencia, entre otras (Zelaya-Molina et al., 2022).
En la naturaleza existen alternativas biológicas basadas en el uso de enemigos naturales los cuales son utilizados en los programas de control biológico validándose como una estrategia rentable y sostenible (Corrales Paredes, 2021). En este contexto, dentro del grupo de los insectos con reconocida actividad biorreguladora se encuentran los coccinélidos, destacados por su actividad depredadora de muchas plagas agrícolas en cultivos de importancia económica (Cevallos Cevallos et al., 2021; Taranto et al., 2022). Algunos de los avances investigativos relacionados al control biológico se han podido desarrollar en cultivos como: chile dulce (Rocca et al., 2021), col (Askar, 2021), nopal (Ascencio Contreras, 2021), algodón (Geethu et al., 2022), soja (Gesraha y Ebeid, 2022) y otros (Rondoni et al., 2021).
En Ecuador, han sido muy limitados los estudios de control biológico, especialmente en el cultivo de maíz, razón por la cual es necesario el conocimiento de los enemigos naturales y, en este contexto, los coccinélidos depredadores. Esta investigación tuvo por objetivo realizar un análisis prospectivo de los coccinélidos asociados al cultivo de maíz en el cantón Santa Ana, provincia de Manabí, Ecuador.
2. Materiales y Métodos
2.1 Ubicación y caracterización del lugar de estudio
La presente investigación se realizó a final de la época seca del año 2015-2016, en los predios de la Facultad de Ingeniería Agronómica de la Universidad Técnica de Manabí [UTM], parroquia Lodana, cantón Santa Ana, provincia de Manabí, Ecuador, localizada a 01°09ʼ51ʼʼS latitud y 80°23ʼ24ʼʼW de longitud, con una altitud de 60 m s.n.m.
2.2 Evaluación de los coccinélidos asociados al cultivo de maíz
El área de estudio utilizada fue de 2.000 m., se emplearon semillas de maíz blanco variedad INIAP-528 con una densidad de siembra equivalente a 40.000 plantas ha-1. El lote estuvo dividido en dos parcelas, una de ellas sujeta a tratamiento químico.
Durante la fase de crecimiento lento (25 dds) surgieron altas poblaciones de la chicharrita (Dalbulus maidis DeLong) la cual fue controlada con el insecticida CETAMISEN® SP (Acetamiprid 250 g de i.a kg-1) a razón de 15 g de producto comercial diluido en 20 L de agua, el cual fue aplicado durante las primeras horas de la mañana (7:30 am) y en una sola ocasión.
No fue necesario hacer uso de otros insecticidas ya que las plagas presentadas se mantuvieron en poblaciones muy bajas y no superaron el umbral económico, excepto D. maidis, cuyo umbral fue mayor a 0,7 individuos, de acuerdo con el promedio establecido por Anasac (2023).
Los insectos colectados fueron adultos, para lo que se emplearon cinco técnicas de muestreo: trampas amarillas, trampas de succión, redada, trampas de luz y las trampas de caída “Pitfall”. Se colocaron cinco trampas amarillas por parcela de estudio con medidas de 0,76 m x 1,25 m, ubicadas en cinco puntos distintos del terreno (“X”) de manera permanente y evitando el efecto borde. A cada una se le colocó grasa vegetal para permitir la adherencia de los insectos.
Las trampas de succión consistieron en colectores plásticos en forma de botella con medidas de 12 cm x 4,5 cm. Su extremo superior estuvo provisto de dos tubos; uno corto con dimensiones de 26 cm x 0,5 cm (obstruido con malla), y, el otro largo con medidas de 36,5 cm x 0,5 cm los cuales permitieron aspirar y capturar al insecto, incluso aquellos localizados en lugares de difícil acceso de la planta. Cada parcela fue muestreada caminando en zig-zag, considerando 5 plantas por sitio de muestreo, es decir, que en cada parcela se evaluaron 25 plantas.
Se realizaron muestreos por medio de la red entomológica; esta herramienta estuvo provista de un mango metálico y una malla muy fina para atrapar los artrópodos. Se evaluaron diez filas de plantas realizando movimientos en forma de “8”. Este tipo de evaluación se efectuó a partir de los 15 días de emergencia del cultivo hasta etapa de crecimiento lento (4.semana) con el fin de evitar daños mecánicos.
Se utilizaron trampas de luz de 15 cm x 10 cm por parcela de estudio. Cada una situada en el centro y provista de una fotocélula de 1,2 v 800 mAh que se recargaba con energía solar durante el día y proyectaba luz ultravioleta en la noche, además, tenía un corto revestimiento de alambre que emitía leves descargas de electricidad. Posterior a esta se le adaptó un cono metálico con medidas de 35 cm de largo y con la apertura superior e inferior de 28 y 7 cm, respectivamente, el cual estuvo sujeto a un recipiente (14 cm x 7,5 cm) que servía para atrapar a los insectos.
Las trampas de caída “Pitfall” consistieron en recipientes de polietileno (botella) con su extremo superior invertido a manera de cono, utilizándose cinco unidades por cada parcela. Para instalarlas, se realizaron hoyos en el suelo, los cuales coincidían con el tamaño de la trampa (14 cm x 7,5 cm), luego fueron colocadas a nivel de la superficie y en cada una se depositó 300 mL de atrayente natural (solución madre formada por 2.700 mL de agua y 300 mL de melaza). La mezcla fue renovada cada semana de muestreo.
La evaluación y recolección de las muestras correspondientes a cada método de trampeo se efectuaron cada 7 días y a un determinado horario (7:30 a.m.) durante ocho semanas consecutivas. Posteriormente fueron trasladadas a un espacio físico adecuado para ser despojadas de impurezas, separadas de otros artrópodos no objetivos de la investigación y conservadas en alcohol al 90 %. Finalmente, se realizó el montaje y se las colocó en sus respectivas cajas entomológicas.
La información obtenida cada semana fue registrada en una matriz, los valores de cada fecha de colecta fueron sumados al final para cada parcela de estudio; consecutivamente, se aplicaron las fórmulas respectivas descritas más adelante.
2.3 Clasificación taxonómica
Los insectos fueron clasificados taxonómicamente en el Laboratorio de Entomología-UTM. Para facilitar su estudio se emplearon materiales bibliográficos de los autores; Iannacone y Perla (2011), Leeper (2016), Bustamante-Navarrete (2020). Bases de datos como: Centro de Biociencia Agrícola Internacional (CABI, 2021), la Colección de Invertebrados del Museo de Zoología QCAZ de la Pontificia Universidad Católica del Ecuador (PUCE, 2021) y la Colección de Insectos del Museo de Entomología de la Universidad del Valle (Posso Gomez, 2013). También se utilizaron las colecciones sinópticas disponibles en la página web “Coccinellidae del Ecuador” (González, 2015). Las muestras fueron corroboradas por la Universidad Nacional Agraria La Molina de Perú.
2.4 Análisis de datos
Los datos fueron analizados por medio de estadística descriptiva y se calcularon índices de diversidad alfa y beta.
Para la determinación del número de individuos, la abundancia relativa y las estimaciones porcentuales, en primer lugar, se realizó la sumatoria de cada una de las especies presentes por cada parcela de estudio, luego se calculó la abundancia relativa [AR] donde se dividió el valor obtenido correspondiente a los individuos de cada especie entre el total de individuos de todas las especies de la parcela de estudio multiplicado por 100 (ecuación [1]). También se obtuvo las estimaciones porcentuales de todas las especies registradas en el área para lograr una visualización generalizada.
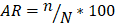 [1]
[1]El índice de riqueza de Margalef [DMg] se calculó mediante la ecuación [2]. Donde S representa el número de especies encontradas, ln el logaritmo natural y . el total de individuos presentes en el sitio.
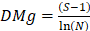 [2]
[2]El índice de Simpson 1-D (dominancia) [λ] se calculó mediante la ecuación [3]. Donde ∑ representa la sumatoria de los individuos, pi el número de individuos de la especie “i” dividido entre el número total de individuos de la muestra (pi = ni/N).
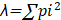 [3]
[3]El Índice de Shannon-Wiener [H’] se calculó mediante la ecuación [4]. Donde pi corresponde a la abundancia relativa, la cual consiste en la división del número de individuos de cada especie entre el total de individuos de todas las especies de la parcela de estudio multiplicado por 100; mientras que ln representa el logaritmo natural y ∑ la sumatoria.
 [4]
[4]El Índice de equitabilidad de Pielou [J] se calculó mediante la ecuación [5]. Donde H’ máx = ln(S), es decir, que H’ corresponde al Índice de Shannon-Wiener, H’máx representa a la diversidad máxima esperada, ln el logaritmo natural y, S constituye el número de especies encontradas.
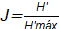 [5]
[5]El índice de Jaccard [IJ] se calculó mediante la ecuación [6]. Donde A representa el número de especies presentes en el sitio A; B el número de especies presentes en el sitio B y C el número de especies comunes presentes en ambos sitios A y B.
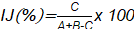 [6]
[6]El Índice de Sorensen [IS] se calculó mediante la ecuación [7]. Donde . representa el número de especies encontradas en la comunidad A; B el número de especies encontradas en la comunidad B y C el número de especies comunes en ambas localidades A y B.
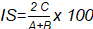 [7]
[7]3. Resultados y Discusión
Se encontraron 297 individuos (Coleoptera: Coccinellidae) distribuidos en 5 subfamilias (Coccinellinae, Coccidulinae, Epilachninae, Ortaliinae y Scymninae) determinándose 13 géneros y 15 especies (Figura 1). La mayor abundancia corresponde a las siguientes especies: Cheilomenes sexmaculata Fabricius con 93 individuos (31,31 %), seguido de Hyperaspis festiva Mulsant con 63 (21,21 %) y Cycloneda sanguinea Linnaeus con 36 individuos (12,12 %) (Tabla 1).

Figura 1
a) Cheilomenes sexmaculata. b) Hyperaspis onerata. c) Hyperaspis festiva. d) Hyperaspis esmeraldas. e) Cyrea alma. f) Psyllobora confluens. g) Hippodamia convergens. h) Tenuisvalvae bromelicola. i) Diomus apollonia. j) Rodolia cardinalis. k) Mada synemia. l) Azya orbigera ecuadorica. m) Cycloneda sanguinea. n) Paraneda pallidula guticollis. o) Scymnus cerinotum.
Figure 1. a) Cheilomenes sexmaculata. b) Hyperaspis onerata. c) Hyperaspis festiva. d) Hyperaspis esmeraldas. e) Cyrea alma. f) Psyllobora confluens. g) Hippodamia convergens. h) Tenuisvalvae bromelicola. i) Diomus apollonia. j) Rodolia cardinalis. k) Mada synemia. l) Azya orbigera ecuadorica. m) Cycloneda sanguinea. n) Paraneda pallidula guticollis. o) Scymnus cerinotum.
| Subfamilia | Especie | N.° de individuos* | Abundancia relativa (%)* | |||||
| SI | CI | Total | SI | CI | Total | |||
| Coccinellinae | Cheilomenes sexmaculata Fabricius | 64 | 29 | 93 | 31,37 | 31,18 | 31,31 | |
| Hippodamia convergens Guérin-Méneville | 15 | 3 | 18 | 7,35 | 3,23 | 6,06 | ||
| Psyllobora confluens Fabricius | 5 | 2 | 7 | 2,45 | 2,15 | 2,36 | ||
| Cycloneda sanguinea Linnaeus | 26 | 10 | 36 | 12,75 | 10,75 | 12,12 | ||
| Paraneda pallidula guticollis Mulsant | 1 | 1 | 2 | 0,49 | 1,08 | 0,67 | ||
| Coccidulinae | Azya orbigera ecuadorica Gordon | 0 | 1 | 1 | 0,00 | 1,08 | 0,34 | |
| Epilachninae | Mada synemia Gordon | 7 | 6 | 13 | 3,43 | 6,45 | 4,38 | |
| Ortaliinae | Rodolia cardinalis Mulsant | 1 | 0 | 1 | 0,49 | 0,00 | 0,34 | |
| Scymninae | Tenuisvalvae bromelicola Sicard | 14 | 8 | 22 | 6,86 | 8,60 | 7,41 | |
| Hyperaspis onerata Mulsant | 3 | 3 | 6 | 1,47 | 3,23 | 2,02 | ||
| Hyperaspis esmeraldas Gordon y González | 2 | 0 | 2 | 0,98 | 0,00 | 0,67 | ||
| Hyperaspis festiva Mulsant | 38 | 25 | 63 | 18,63 | 26,88 | 21,21 | ||
| Scymnus cerinotum Gordon | 11 | 2 | 13 | 5,39 | 2,15 | 4,38 | ||
| Cyrea alma Gordon & Canepari | 16 | 3 | 19 | 7,84 | 3,23 | 6,40 | ||
| Diomus apollonia Gordon | 1 | 0 | 1 | 0,49 | 0,00 | 0,34 | ||
| Sumatoria | 204 | 93 | 297 | 100 | 100 | 100 | ||
Table 1. Number of individuals, relative abundance and percentage estimation of coccinellids associated with maize crop, at the “La Teodomira” experimental campus, Santa Ana, Ecuador.
* SI: Sin aplicación de Insecticida. CI: Con aplicación de insecticida. / SI: Without insecticide application; CI: With insecticide application.En la parcela sin aplicación de insecticida [SI] se observaron 204 individuos, distribuidos en 12 géneros y 14 especies, destacándose Ch. sexmaculata con 64 individuos (31,37 %), H. festiva con 38 (18,63 %) y C. sanguinea con 26 individuos (12,75 %) (Tabla 1). En la parcela con aplicación de insecticida [CI] se encontraron 93 individuos, 11 géneros y 12 especies, sobresaliendo Ch. sexmaculata con 29 individuos (31,18 %), H. festiva con 25 (26,88 %) y C. sanguinea con 10 individuos (10,75 %) (Tabla 1).
Los resultados mostraron menor abundancia de coccinélidos en la parcela tratada con insecticida. Cardoso De Sousa et al. (2021) y Skouras et al. (2022) sostienen que la familia Coccinellidae se encuentra estrictamente afectada por las aplicaciones de agroquímicos, los cuales atentan contra la supervivencia de estos depredadores generalistas. En este sentido, las “mariquitas” se encuentran expuestas a los insecticidas por medio de tres vías distintas; contacto directo a superficie tratadas, ingestión de presas contaminadas y por contacto residual, al que las larvas y los huevos pueden estar expuestos de forma persistente. En correspondencia a lo mencionado existen algunos estudios detallados a continuación que argumentan la toxicidad de los insecticidas respecto a este tipo de controlador biológico:
Cheng et al. (2022) determinaron la toxicidad y evaluación de los riesgos de nueve pesticidas en depredadores naturales, no objetivos de Harmonia axyridis, donde se utilizaron siete insecticidas (imidacloprid, dinotefuran, tiametoxam, acetamiprid, bifentrina, dimetoato y emamectin benzoato) y dos fungicidas (tebuconazole y myclobutanil). En el estudio se constató que casi todos los insecticidas utilizados representaron alto riesgo de mortalidad para el coccinélido, a excepción del emamectin benzoato y los dos fungicidas, los cuales reportaron menor riesgo para el depredador.
Dai et al. (2021) evaluaron los efectos transgeneracionales del imidacloprid y el insecticida sulfoximina en la mariquita alerquín (H. axyridis). Los resultados mostraron que las tasas de eclosión, supervivencia y emergencia de las larvas disminuyeron significativamente, y que las etapas de huevo y estadio de la generación F1 aparentemente podrían prolongarse cuando su generación parental F0 estuvo expuesta a dichos insecticidas en concentraciones de LC 20 y LC 50. Por lo tanto, sugiere que se usen con precaución en los programas de MIP contra A. gossypii, de lo contrario, se reducirá el servicio ecológico y la eficacia de las mariquitas arlequín.
Similar resultado obtuvo Pavithrakumar et al. (2022) quienes evaluaron la toxicidad de algunos insecticidas sobre los estadios de Ch. sexmaculata bajo condiciones de laboratorio. En los bioensayos de toxicidad aguda realizados a las dosis de campo recomendadas mostraron que el dimetoato era dañino para todas las etapas de la vida con un 100 % de mortalidad. También se encontró que el tiametoxam era seguro hasta la etapa de huevo; sin embargo, fue dañino para las larvas, las pupas y los adultos. Mientras que el aceite de neem fue perjudicial para las etapas no alimentarias (huevos y pupas), inhibiendo completamente la eclosión de huevos y la emergencia de adultos; además, fue inofensivo o levemente dañino para las etapas alimentarias. Por otro lado, se constató que la flubendiamida y el spinosad presentaron una mortalidad > 10 % en huevos, larvas y pupas y 12,5 % para adultos. También existen otras contribuciones donde se evalúa el efecto de los insecticidas, cuyo propósito es garantizar aquellos que puedan tener mayor control de plagas y menor impacto sobre los insectos benéficos, en este contexto, se apoya en las investigaciones desarrolladas por Barbosa et al. (2018), da Silva et al. (2022), Skouras et al. (2022) y You et al. (2022).
Con relación a las especies destacadas en el estudio (Ch. sexmaculata, H. festiva y .. sanguinea) se atribuyó que estas podrían encontrarse favorecidas por las condiciones ambientales del lugar, tales como: temperatura media anual de 27,7 .C, precipitación anual acumulada de 544 mm y humedad relativa que oscila entre 70-80 %. Otro factor importante es la vegetación circundante, por ejemplo, se observaron cultivos de ciclo corto (solanáceas y leguminosas), cultivos perennes como cacao y cítricos, también fue común encontrar especies arvenses como principales hospederas de áfidos y mosca blanca, constituyendo presas para garantizar la supervivencia y abundancia de los coccinélidos. Los resultados de esta investigación coinciden con los estudios realizados por Sruthi et al. (2021), Martínez Chiguachi (2018) y Erráez Aguilera et al. (2020), quienes reportaron mayor abundancia de estas especies en cultivos variados (berenjena, caupí, ají, cítricos y otros cultivos).
3.1 Diversidad alfa y beta
El índice de Margalef se centra en la riqueza de las especies que se encuentran en una población determinada. En cada parcela se obtuvieron valores correspondientes a 2,44 (SI) y 2,43 (CI) con 14 y 12 especies, respectivamente. Según Duarte-Goyes et al. (2019), estos valores están comprendidos dentro de los rangos 2 y 2,7, interpretándose como agroecosistemas de diversidad media. Martínez Chiguachi (2018) sostiene que esta condición es atribuida a factores antrópicos, tales como las aplicaciones de pesticidas, falta de rotación, cultivos asociados, etc.
El índice de Simpson permite conocer la probabilidad de que dos individuos de una muestra comunitaria pertenezcan a la misma especie (Wu et al., 2022). Los valores encontrados en esta investigación (0,82 SI y 0,80 CI), sugieren alta diversidad e indican baja dominancia (0,17 SI y 0,19 CI), al ser un índice inversamente proporcional de acuerdo con la escala mencionada por Kunda et al. (2022) cuyo rango de interpretación varía entre 0 y 1. Estudios similares se han encontrado con el trabajo realizado por Cisneros Puma (2021), quien evaluó la diversidad y abundancia de los coccinélidos asociados a los cultivos de alfalfa y maíz en la provincia de Arequipa Perú y comprobó mayor dominancia de especies en ambos cultivos durante la época lluviosa en relación a la época seca, condición atribuida principalmente a los factores climáticos. Resultados opuestos se han podido obtener con la investigación de Cuello Catalán (2022) quien estudió la variación interanual de coccinélidos nativos y exóticos en la última década y su relación con los cambios climáticos en la región metropolitana de Santiago de Chile. Comprobó mayor diversidad durante los años 2010 y 2011; sin embargo, en el 2012 hasta 2021 se evidenció niveles de diversidad significativamente bajos, debido al cambio climático, el cual amenaza la biodiversidad, en conjunto con la contaminación, destrucción de hábitats, invasiones biológicas y otros factores antropogénicos.
El índice de Shannon-Wiener basado en la riqueza de especies y la abundancia proporcional reportó en las parcelas estudiadas valores calculados de 2,06 (SI) y 1,94 (CI) los cuales se encuentran dentro de los rangos comprendidos entre 1,5 y 3,5 de acuerdo con lo propuesto por Kunda et al. (2022), quienes lo asocian con agroecosistemas de diversidad media. Esta información concuerda con la investigación desarrollada por Rasheed y Buhroo (2018), quienes estudiaron la diversidad de escarabajos coccinélidos en tres distritos del sur de Cachemira, India, demostrando valores de 2,29, 2,30 y 2,33, determinándose agroecosistemas de diversidad media entre los sitios estudiados.
El índice de Pielou demostró un comportamiento equitativo de las especies presentes en la investigación, con resultados de 0,78 para cada parcela estudiada (Tabla 2), lo cual estuvo dentro del rango referido por Kunda et al. (2022) quienes sugieren una escala que oscila entre 0 y 1, los resultados cercanos a 1 indican comunidades equitativas.
| Parámetros | Parcelas* | |
| SI | CI | |
| Número total de individuos (N) | 204 | 93 |
| Número total de especies (S) | 14 | 12 |
| Índice de Margalef | 2,44 | 2,43 |
| Índice de Shannon | 2,06 | 1,94 |
| Diversidad de Simpson | 0,82 | 0,80 |
| Dominancia de Simpson | 0,17 | 0,19 |
| Índice de Pielou | 0,78 | 0,78 |
| Índice de Jaccard (%) | 73,33 | |
| Índice de Sorensen (%) | 84,62 | |
Table 2. Alpha and beta diversity of coccinellids associated with maize crop, at the “La Teodomira” experimental campus, Santa Ana, Ecuador.
* SI: Sin aplicación de insecticida; CI: Con aplicación de Insecticida. / SI: Without insecticide application; CI: With insecticide application.Respecto a los índices de similaridad de Jaccard y Sorensen, demostraron que en ambas parcelas existe alta similitud de especies encontradas, reportando valores de 73,33% (I. J) y 84,62% (I. S), Ramírez et al. (2015) sustentan que los valores aproximados a 1 indican alta similaridad. Esta investigación coincide con el estudio desarrollado por Cisneros Puma (2021), quien encontró en el cultivo de alfalfa alta similaridad de coccinélidos, reportando valores del 78 y 88 % mediante el índice de Jaccard y Sorensen, respectivamente, en las localidades de Cayma y Polobaya, Perú (época seca), mientras que en la época lluviosa alcanzaron valores del 45 % (I. J) y 63 % (I. S).
3.2 Abundancia de coccinélidos de acuerdo con las técnicas de muestreo
Se determinó que la técnica de muestreo que capturó el mayor número de coccinélidos fueron las trampas cromáticas (amarillas), con un total de 275 individuos, seguidos de la trampa de succión y la redada. Estas constituyen estrategias importantes para el monitoreo de artrópodos, no obstante, su eficiencia dependerá del tipo de artrópodo que se quiera capturar, tipo de trampa, color a utilizar, altura, tipo de cultivo, entre otras. Ikemoto et al. (2021) evaluaron la eficiencia de diferentes tipos de trampas para la atracción de tres órdenes de insectos (Hymenoptera, Diptera y Coleoptera) y comprobaron que la mayor abundancia de coleópteros se presentó en las trampas adhesivas amarillas, comprobando alta efectividad; en este sentido, las reacciones de los insectos al color son generadas por sus sistemas visuales, características ecológicas como hábitat y alimento. Los insectos fitófagos y sus depredadores o parasitoides generalmente se sienten atraídos por el color amarillo, probablemente porque este color representa un estímulo sobresaliente relacionado con el follaje.
Por otro lado, Mahalakshmi et al. (2022) manifiestan que las trampas adhesivas amarillas pueden llegar a ser beneficiosas para la captura de plagas, pero perjudiciales para los insectos no objetivos, tales como los coccinélidos, razón por la cual deberían emplearse con precaución, salvo el caso de implementar esta técnica como medida para generar mayor conocimiento ecológico sobre las especies depredadoras (Tabla 3).
| Trampas | SI* | CI* | Total |
| Adhesivas Amarillas | 195 | 80 | 275 |
| Redada | 1 | 7 | 8 |
| Succión | 8 | 6 | 14 |
| Pitfall | 0 | 0 | 0 |
| Luz | 0 | 0 | 0 |
| Sumatoria | 204 | 93 | 297 |
Table 3. Traps used to capture coccinellids associated with maize crop, at the “La Teodomira” experimental campus, Santa Ana, Ecuador.
* SI: Sin aplicación de insecticida; CI: Con aplicación de Insecticida. / SI: Without insecticide application; CI: With insecticide application.3.3 Aspectos biológicos de los principales coccinélidos registrados en el estudio
Según Merlin et al. (2022), el potencial de alimentación y reproducción de los depredadores se encuentra influenciado por las plantas debido al efecto directo sobre la calidad de sus presas y también por las condiciones ambientales, las cuales inciden sobre su ciclo de vida, representando un factor determinante para su cría masiva. Rodríguez et al. (2019) relatan que los parámetros biológicos y reproductivos de los enemigos naturales son atributos importantes que dan pautas para ser considerados dentro de los programas de control biológico. A continuación, se mencionan algunos aspectos relacionados al ciclo de vida de las principales especies destacadas en el estudio:
3.3.1 Cheilomenes sexmaculata
Ashwini y Shukla (2022) evaluaron el ciclo de vida de Ch. sexmaculata alimentados con el pulgón Aphis craccivora(Koch) bajo condiciones de laboratorio a 27,18 ± 1,45 ℃ y 49,80 ± 7,64 % de humedad relativa. Los resultados revelaron que el período medio de incubación de dicho depredador fue de 1,64±0,72 días, el porcentaje medio de eclosión de los huevos fue 89,25±3,97 %, la duración media del primer, segundo, tercer y cuarto estadio larvario fue de 1,26±0,44; 1,68±0,59; 2,28±0,54 y 2,44±0,61 días, respectivamente. El período larvario total fue de 7,66±1,61 días. El período medio prepupal y pupal fue de 1,04±0,20 y 2,18±0,48 días, independientemente. El porcentaje medio de emergencia de adultos fue de 89,48±4.44 % con una proporción de sexos de 1:1,40. La pre-oviposición media, oviposición y post-oviposición fue de 3,48±0,77, 16,40±1,29 y 3,00±0,87 días, respectivamente. La fecundidad de una hembra grávida fue de 363,32±76,32 huevos. La longevidad en adultos machos y hembras fue 17,12±1,44 días y 22,54±2,38 días, respectivamente. La esperanza de vida total de los machos fue de 29,64±2,59 mientras que en las hembras fue de 35,06±3,17 días.
Existen otras contribuciones como la de Abbas et al. (2020) quienes evaluaron el ciclo biológico de C. sexmaculata bajo condiciones controladas (25 ± 2 ⁰C y 60 ± 5 % humedad relativa) el cual fue alimentado con cuatro especies de áfidos (Lipaphis erysimi Kaltenbach, Myzus persicae Sulzer, Aphis nerii Boyer de Fonscolombe y Diuraphis noxia Mordvilko) y comprobaron que el huésped más apropiado para la alimentación del coccinélido fue D. noxia. En este contexto, detalló la información obtenida acerca de los parámetros biológicos y reproductivos: la duración de la etapa de huevos fue de 3,30±0,16 días; el primer instar conllevó de 1,00±2,19 días; el segundo instar fue de 1,10±0,7 días; el tercer instar fue de 1,47±1,12 días; el cuarto instar fue de 2,47±0,12 días; la pupa presentó una duración de 2,65±3,48 días; la fase de huevo hasta adulto fue de 12,42±0,35 días; la longevidad del macho y la hembra fue de 26,33±2,19 y 28,00±0,83 días, respectivamente; la pre-oviposición, oviposición y post-oviposición fue de 5,33±1,33; 14,70±1,33 y 7,33±1,70 días, independientemente, además, alcanzó una fecundidad 316,80±25,07 huevos por hembra.
3.3.2 Hyperaspis spp.
Ascencio Contreras (2021) determinó la duración promedio de los estados biológicos de cuatro coccinélidos alimentados con hembras adultas de la grana silvestre (Dactylopius opuntiae Cockerell). Dentro del complejo de enemigos naturales evaluó a Hyperaspis trifurcata Schaeffer bajo condiciones controladas (28 ± 2 °C y 50 % de humedad relativa) dando a conocer los siguientes parámetros biológicos; fase de huevo 5,41 días, larva I-IV 16,48 días, pre pupa 1,33 días, pupa 7 días, adulto 39,5 días.
3.3.3 Cycloneda sanguinea
Rodríguez et al. (2019) evaluaron los aspectos biológicos y parámetros poblacionales de .. sanguinea alimentado con el pulgón Aphis aurantii Boyer de Fonscolombe registrando a nivel de laboratorio (25 ± 2 °C y 70 ± 10 % de humedad relativa) la siguiente longevidad; la fase de huevo alcanzó 3,78 días, el desarrollo larvario I-IV 10,34 días, la pre pupa 1,74 días, la pupa 3,46 días, los adultos machos y hembras 58,37 y 61,71 días, respectivamente, la pre-oviposición tardó 3,9 días, donde cada hembra colocó durante toda su vida 882,2 huevos alcanzando una tasa de oviposición de 34,6 huevos/hembra/día. La duración promedio de la fase de huevo-adulto fue de 19,32 días.
Santos-Cividanes et al. (2022) evaluaron la supervivencia y fecundidad de C. sanguinea alimentadas con el chinche verde Schizaphis graminum Rondani bajo condiciones controladas (25 ± 1 °C y 70 ± 10 % de humedad relativa) y determinó que las hembras colocaron huevos entre los 3-4 días posterior a la adultez, el período de oviposición se extendió desde el día 18 hasta el 105 y alcanzó su cúspide en el día 24 con un total de 17,6 huevos por hembra. Consecutivamente, la tasa de supervivencia de adultos disminuyó gradualmente a partir del día 33 y alcanzaron la supervivencia máxima en el día 92.
Rodríguez-Palomera et al. (2015) argumentan que los parámetros biológicos y reproductivos pueden variar en función del tipo de alimento al que están expuestos los coccinélidos, conjuntamente con los factores ambientales, lo que implica que podría alargarse o acortarse su ciclo de vida. Islam et al. (2022) y Su et al. (2023) sustentan que cuando el alimento ingerido es escaso o de menor calidad el tiempo de desarrollo suele aumentar y la tasa reproductiva disminuye.
4. Conclusiones
Se determinó que los insectos benéficos procedentes de la familia Coccinellidae, son sensibles ante la aplicación de agroquímicos, lo que repercute en su abundancia y diversidad.
El índice de Margalef obtenido en las parcelas de estudio fue de 2,44 (SI) y 2,43 (CI) con 14 y 12 especies, respectivamente. Estos valores están comprendidos dentro de los rangos 2 y 2,7 interpretándose como agroecosistemas de diversidad media.
Se constató que los índices de Simpson encontrados en esta investigación (0,82 SI y 0,80 CI), sugieren alta diversidad y baja dominancia (0,17 SI y 0,19 CI).
El índice de Shannon-Wiener basado en la riqueza de especies y la abundancia proporcional reportó en las parcelas estudiadas valores calculados de 2,06 (SI) y 1,94 (CI) los cuales se encuentran dentro de los rangos comprendidos entre 1,5 y 3,5 catalogándose como un agroecosistema de diversidad media.
El índice de Pielou, demostró que las comunidades presentaron un comportamiento equitativo con un valor de 0,78 para cada parcela estudiada ya que el índice se aproxima a 1.
Respecto a los índices de similaridad de Jaccard y Sorensen, demostraron que en ambas parcelas existe alta similitud de especies encontradas, reportando valores de 73,33 % (I. J) y 84,62 % (I. S), dado que los valores se aproximan a 1.
La técnica más eficiente para la captura de coccinélidos fue la trampa cromática lo que representa una ventaja para efectuar estudios fundamentados en el conocimiento de este tipo de depredador, no obstante, se recomendaría utilizarlo con precaución ya que este método de trampeo usualmente es utilizado para captura de plagas en campo, razón por la cual sería importante considerar otros criterios como el color a utilizar, tipo de plagas que se requiere capturar o disminuir su población de campo, tipo de cultivo, dimensiones de la trampa, entre otros parámetros, los cuales permitan minimizar la atracción de los insectos benéficos.
Contribuciones de los autores
-
Jessica Daniela Zambrano Mero: recursos, conceptualización, investigación, metodología, redacción – borrador original.
-
Nexar Emiliano Vega Lucas: recursos, redacción – revisión y edición.
-
Ariolfo Leonardo Solís Bowen: supervisión, redacción – revisión y edición.
-
Dorys Terezinha Chirinos Torres: validación, redacción – revisión y edición.
-
Diego Rodolfo Perla Gutiérrez: Investigación (procesamiento taxonómico de insectos).
-
Alex Gabriel Delgado Párraga: curación de datos.
-
Sofía Lorena Peñaherrera Villafuerte: análisis formal
Implicaciones éticas
Los autores declaran que no existen implicaciones éticas.
Conflicto de interés
Los autores declaran que no existen conflictos de interés financieros o no financieros que podrían haber influido en el trabajo presentado en este artículo.
Agradecimientos
Se agradece a la Universidad Técnica de Manabí [UTM], a la Universidad Nacional Agraria La Molina [UNALM] y a los coautores del presente artículo por colaboración brindada y cumplimiento de actividades bajo rigor científico. A la Revista “SIEMBRA” por hacer posible la difusión de los resultados.
Referencias
Abbas, K., Zaib, M. S., Zakria, M., Hani, U., Zaka, S. M, y Ane, M. (2020). Cheilomenes sexmaculata (Coccinellidae: Coleoptera) as a potential biocontrol agent for aphids based on age-stage, two-sex life table. PLOS ONE, 15(9), e0228367. https://doi.org/10.1371/journal.pone.0228367
Anasac. (2023). Aprendamos de Dalbulus maidis. Boletín técnico I + D. https://anasac.co/boletin-tecnico-agro_-dalbulus-maidis/
Ascencio Contreras, D. O. (2021). Coccinélidos como enemigos naturales de Dactylopius opuntiae (Cockerell). Universidad Autónoma de Nuevo León. http://eprints.uanl.mx/id/eprint/20997
Askar, S. I. (2021). Efficiency of three coccinellid species against Brevicoryne brassicae (L.) (Homoptera: Aphididae) in cabbage fields at El-Behera Governorate, Egypt. Journal of Plant Protection and Pathology, 12(1), 31-35. https://dx.doi.org/10.21608/jppp.2021.149518
Ashwini, M., y Shukla, A. (2022). Biology of zigzag ladybird beetle, Cheilomenes sexmaculata (Fabricius) (Coccinellidae: Coleoptera) on Cowpea aphid, Aphis craccivora (Koch) (Aphididae: Hemiptera). The Pharma Innovation, 11(8), 813-818. https://www.thepharmajournal.com/special-issue?year=2022&vol=11&issue=8S&ArticleId=14834
Barbosa, P. R. R., Oliveira, M. D., Barros, E. M., Michaud, J. P., y Torres, J. B. (2018). Differential impacts of six insecticides on a mealybug and its coccinellid predator. Ecotoxicology and Environmental Safety, 147, 963-971. https://doi.org/10.1016/j.ecoenv.2017.09.021
Barreto Carbajal, J. S., Bustinza Saldívar, D., Álvarez Arias, C., Kari Ferro, A., Huaraca Aparco, R., Flores Pacheco, N. F., y Echegaray Peña, N. G. (2022). Contaminantes agrícolas en la producción de maíz Zea mays L. variedad choclera en Curahuasi Apurimac, Perú. Revista de Investigación en Ciencia y Tecnología: C&T Riqchary, 4(1), 20-25. https://doi.org/10.57166/riqchary/v4.n1.2022.84
Bustamante-Navarrete, A. A. (2020). Algunos coccinellidos (Coleóptera: Coccinellidae) predadores de importancia económica en el departamento del Cusco, Perú. The Biologist, 18(2), 287-314. https://doi.org/10.24039/rtb2020182801
CABI. (2021). CABI Compendium Invasive Species. https://www.cabidigitallibrary.org/product/QI
Cardoso De Sousa, M., Andaló, V., Zampiroli, R., Batista De Alvarenga, C., Aparecida De Assis, G., Janoni Carvalho, F., De Brito, W. A., y Silva De Faria, L. (2021). Impact of a synthetic insecticide and plant extracts on Aphis craccivora (Hemiptera: Aphididae) and coccinellid species in common beans. Revista Colombiana de Entomología, 47(2). https://doi.org/10.25100/socolen.v47i2.10992
Cevallos Cevallos, D., Santana Cedeño, J., y Chirinos, D. T. (2021). Los depredadores y el manejo de algunas plagas agrícolas en Ecuador. Manglar, 18(1), 51-59. http://dx.doi.org/10.17268/manglar.2021.007
Cheng, S., Yu, C., Xue, M., Wang, X., Chen, L., Nie, D., Zhang, N., Zhang, J., Hou, Y., y Lin, R. (2022). Toxicity and risk assessment of nine pesticides on nontarget natural predator Harmonia axyridis (Coleoptera: Coccinellidae). Pest Management Science, 78(12), 5124-5132. https://doi.org/10.1002/ps.7130
Cisneros Puma, L. A. (2021). Diversidad y abundancia de la familia Coccinellidae asociados a cultivos de alfalfa y maíz de los distritos de Cayma y Polobaya, provincia de Arequipa - 2019. Universidad Nacional de San Agustín de Arequipa. http://hdl.handle.net/20.500.12773/12446
Corrales Paredes, C. J. (2021). Uso de depredadores, parasitoides y entomopatógenos para el control biológico de plagas y enfermedades de hortalizas con énfasis en tomate (Solanum lycopersicum L.). Universidad Técnica de Cotopaxi. https://repositorio.utc.edu.ec/handle/27000/7622
Cuello Catalán, K. A. (2022). Variación interanual de coccinélidos nativos y exóticos presentes en cultivos de alfalfa en la última década y su relación con cambios climáticos en la región Metropolitana de Santiago. Universidad de Chile. https://repositorio.uchile.cl/handle/2250/194575
Dai, C., Ricupero, M., Wang, Z., Desneux, N., Biondi, A., y Lu, Y. (2021). Transgenerational effects of a neonicotinoid and a novel sulfoximine insecticide on the harlequin ladybird. Insects, 12(8), 681. https://doi.org/10.3390/insects12080681
Duarte-Goyes, A. M., Rosero-Erazo, L. F., Guerra-Acosta, A. del S., y Moreno Ortega, P. E. (2019). Diversidad biológica y seguridad alimentaria que ofrecen las huertas urbanas del altiplano andino – amazónico en el Valle de Sibundoy, Putumayo, Colombia. Cuadernos de Biodiversidad, (57), 12-25. https://doi.org/10.14198/cdbio.2019.57.02
Erenstein, O., Jaleta, M., Sonder, K., Mottaleb, K., y Prasanna, B. M. (2022). Global maize production, consumption and trade: trends and R&D implications. Food Security, 14, 1295-1319. https://doi.org/10.1007/s12571-022-01288-7
Erráez Aguilera, M., Mazón, M., Troya Armijos, H., y Valarezo Espinoza, D. (2020). Identificación y evaluación de la incidencia de insectos y hongos benéficos asociados a Diaphorina citri Kuwayama (Hemiptera: Liviidae) en plantas traspatio (Citrus spp. y Murraya paniculata) del cantón Catamayo (Loja-Ecuador). Ecuador es calidad, 7(1), 25-33. https://revistaecuadorescalidad.agrocalidad.gob.ec/revistaecuadorescalidad/index.php/revista/article/view/99
FAOSTAT. (2023). Cultivos y producción de ganadería. Food and Agriculture Organization Corporate Statistical Database. https://www.fao.org/faostat/es/#data
Fernández Aravena, C. I. (2021). Alteraciones tiroideas en agricultores de Cotopaxi y su relación con el uso de plaguicidas. Revista San Gregorio, (45), 32-46. https://doi.org/10.36097/rsan.v0i45.1396
Geethu, S., Suroshe, S. S., Chander, S., y Venkanna, Y. (2022). Interactions of six spotted ladybird beetle, Cheilomenes sexmaculata (F.) with its host Phenacoccus solenopsis Tinsley and Intraguild members. International Journal of Tropical Insect Science, 42, 885-893. https://doi.org/10.1007/s42690-021-00614-4
Gesraha, M. A., y Ebeid, A. R. (2022). Releasing two important insect predators to control of aphids under open-field and greenhouse conditions. Asian Journal of Biology, 14(3), 14-23. https://doi.org/10.9734/ajob/2022/v14i330215
González, E. (2015). Los Coccinellidae de Ecuador. https://www.coccinellidae.cl/paginasWebEcu/Paginas/Introduccion_Ecu_01.php
Hernández-Trejo, A., Estrada Drouaillet, B., Rodríguez-Herrera, R., García Giron, J. M., Patiño-Arellano, S. A. A., y Osorio-Hernández, E. (2019). Importancia del control biológico de plagas en maíz (Zea mays L.). Revista Mexicana de Ciencias Agrícolas, 10(4), 803-813. https://doi.org/10.29312/remexca.v10i4.1665
Iannacone, J., y Perla, D. (2011). Invasión del depredador Harmonia axyridis (Coleoptera: Coccinellidae) y una evaluación del riesgo ambiental en el Perú. The Biologist, 9(2), 213-233. https://doi.org/10.24039/rtb201192494
Ikemoto, M., Kuramitsu, K., Sueyoshi, M., Seguchi S., y Yokoi, T. (2021). Relative trapping efficiencies of different types of attraction traps for three insect orders in an agricultural field. Applied Entomology and Zoology, 56, 393-405. https://doi.org/10.1007/s13355-021-00748-4
Instituto Nacional de Estadística y Censos [INEC]. (2023).Estadísticas Agropecuarias. Tabulados de la Encuesta de Superficie y Producción Agropecuaria Continua ESPAC, 2021. INEC. https://www.ecuadorencifras.gob.ec/estadisticas-agropecuarias-2/
Islam, Y., Güncan, A., Zhou, X., Naeem, A., y Shah, F. M. (2022). Effect of temperature on the life cycle of Harmonia axyridis (Pallas), and its predation rate on the Spodoptera litura (Fabricius) eggs. Scientific reports, 12, 15303. https://doi.org/10.1038/s41598-022-18166-z
Kunda, M., Ray, D., Pandit, D., y Harun-Al-Rashid, A. (2022). Establishment of a fish sanctuary for conserving indigenous fishes in the largest freshwater swamp forest of Bangladesh: A community-based management approach. Heliyon, 8(5), e09498. https://doi.org/10.1016/j.heliyon.2022.e09498
Leeper, J. R. (2016). Key to the Coccinelids (Coleoptera: Coccinellidae) established in Hawaii. University of Hawaii e vols series. http://hdl.handle.net/10524/49028
Mahalakshmi, M., Sreekanth, M., Bindu Madhavi, G., y Ramana, M. V. (2022). Influence of traps colour and height of its placement in trapping sucking pests and natural enemies in greengram. Journal of Eco-Friendly Agriculture, 17(1), 137-142. https://doi.org/10.5958/2582-2683.2022.00028.4
Martínez Chiguachi, J. A. (2018). Conservation of non-crop plants as a management strategy to improve functional biodiversity in chili pepper crops. Universidade Federal de Viçosa. https://www.locus.ufv.br/handle/123456789/27263
Merlin, B. L., Ferreira, L. P., Godoy, W. A. C., Moraes, G. J., y Cônsoli, F. L. (2022). Functional response of Neoseiulus californicus preying on Tetranychus urticae is affected by prey quality and host-plant acclimation. Biological Control, 165, 104811. https://doi.org/10.1016/j.biocontrol.2021.104811
Pavithrakumar, K., Smitha, M., Chellappan, M., y Subramanian, M. (2022). Toxicity of some insecticides to lifestages of Cheilomenes sexmaculata under laboratory conditions. Indian Journal of Entomology, 84(4), 878-880. https://doi.org/10.55446/IJE.2022.157
Posso Gomez, C. E. (2013). Base de datos de la colección de insectos del Museo de Entomología de la Universidad del Valle. https://www.gbif.org/es/dataset/67c3b5e0-0f27-4279-97ab-c8f5bf12a97c
Pontificia Universidad Católica del Ecuador [PUCE]. (2021).Base de datos de la colección de invertebrados del Museo de Zoología QCAZ. Versión 2021.0. https://bioweb.puce.edu.ec/QCAZ/baseDatos
Ramírez, J., Hoyos, V., y Plaza, G. (2015). Phytosociology of weeds associated with rice crops in the department of Tolima, Colombia. Agronomía Colombiana, 33(1), 64-73. https://doi.org/10.15446/agron.colomb.v33n1.46747
Rasheed, R., y Buhroo, A. A. (2018). Diversity of coccinellid beetles (Coccinellidae: Coleoptera) in Kashmir, India. Entomon, 43(2), 129-136. https://entomon.in/index.php/Entomon/article/view/375
Rocca, M., Díaz Lucas, M. F., y Greco, N. M. (2021). Effect of spatiotemporal association and trophic interactions between Aphidophagous Coccinellids Toward Aphid Control. Environmental Entomology, 51(1), 44-51. https://doi.org/10.1093/ee/nvab127
Rodríguez-Palomera, M., Cambero-Campos, J., Luna-Esquivel, G., Robles-Bermúdez, A., Yáñez-Celedón, C., y Cambero-Nava, K. G. (2015). Ciclo de vida de Cycloneda sanguinea Linnaeus (Coleoptera: Coccinellidae) sobre el pulgón Aphis aurantii Boyer de Fonscolombe (Hemiptera: Aphididae) en Nayarit, México. En XXXIX Congreso Nacional de Control Biológico. Sociedad Mexicana de Control Biológico. Guadalajara.
Rodríguez-Palomera, M., Luna-Esquivel, G., Robles-Bermúdez, A., Coronado Blanco, J. M., Cambero-Nava, K. G.,.y Cambero-Campos, O. (2019). Aspectos biológicos y parámetros poblacionales de Cycloneda sanguinea L. (Coleoptera: Coccinellidae) sobre Aphis aurantii Boyer de Fonscolombe (Hemiptera: Aphididae). Chilean Journal of Agricultural & Animal Sciences, 35(2), 196-204. https://doi.org/10.4067/S0719-38902019005000305
Rondoni, G., Borges, I., Collatz, J., Conti, E., Costamagna, A. C., Dumont, F., Evans, E. W., Grez, A. A., Howe, A. G., Lucas, E., Maisonhaute, J-E., Soares, A. O., Zavieso, T., y Cock, M. J. W. (2021). Exotic ladybirds for biological control of herbivorous insects - a review. Entomología Experimentalis et Applicata, 169(1), 6-27. https://doi.org/10.1111/eea.12963
Santos-Cividanes, T. M., Cividanes, F. J., Souza, L. S., Matos, S. T. S., y Ramos, T. O. (2022). Life tables of the ladybird beetles Harmonia axyridis, Cycloneda sanguinea and Hippodamia convergens reared on the greenbug Schizaphis graminum. Brazilian Journal of Biology, 82, e263276. https://doi.org/10.1590/1519-6984.263276
Tanumihardjo, S. A., McCulley, L., Roh, R., Lopez-Ridaura, S., Palacios-Rojas, N., y Gunaratna, N. S. (2020). Maize agro-food systems to ensure food and nutrition security in reference to the Sustainable Development Goals. Global Food Security, 25, 100327. https://doi.org/10.1016/j.gfs.2019.100327
da Silva, W. R., Pereira, R. C., Mendonça, L. V. P., Peçanha, L. S., de Sales Abreu, L. M., Abib, P. H. N., Samuels, R. I., Picanço, M. C., y Silva, G. A. (2022). Lethal and sublethal effects of insecticidesused in the management of Plutella xylostella (Lepidoptera: Plutellidae) on the predator Cycloneda sanguinea L. (Coleoptera:Coccinellidae). Pest Management Science, 78(10), 4397-4406. https://doi.org/10.1002/ps.7060
Skouras, P., Demopoulos, V., Mprokaki, M., Anagnostelis, K., Darras, A. I., Stournaras, V., Delis, C., y Stathas, G. J. (2022). Relative toxicity of two insecticides to Coccinella septempunctata and Hippodamia variegata (Coleoptera: Coccinellidae): Implications for integrated management of the aphids, Myzus persicae and Aphis fabae (Hemiptera: Aphididae). Phytoparasitica, 50, 141-150. https://doi.org/10.1007/s12600-021-00935-7
Sruthi, M., Raguraman, S., Chitra, N., y Balaji, R. (2021). Comparative abundance and diversity of predatory coccinellids in bhendi, brinjal, and cowpea in Coimbatore and Tiruppur districts in Tamil Nadu, India. The Pharma Innovation Journal, 10(10S), 1128-1135. https://www.thepharmajournal.com/special-issue?year=2021&vol=10&issue=10S&ArticleId=8531
Su, W., Ouyang, C., Li, Z., Yuan, Y., Yang, Q., y Ge, F. (2023). Cnidium monnieri (L.) Flor de Cusson como alimento complementario que promueve el desarrollo y la reproducción de las mariquitas Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Plants, 12(9), 1786. https://doi.org/10.3390/plants12091786
Taranto, L., Rodríguez, I., Santos, S. A. P., Villa, M., y Pereira, J. A. (2022). Response of the coccinellidae community within sustainable vineyards to the surrounding landscape. Agronomy, 12(9), 1-13. https://doi.org/10.3390/agronomy12092140
Vargas Sunta, I. F., y Moyano Calero, W. (2022). Efectos de los plaguicidas sobre la salud humana en una comunidad de agricultores. Sapienza International Journal of Interdisciplinary Studies, 3(5), 229-238. https://doi.org/10.51798/sijis.v3i6.482
Wu, Y., Cheng, Y., Wang, X., Fan, J., y Gao, Q. (2022). Spatial omics: Navigating to the golden era of cancer research. Clinical and Translational Medicine, 12(1), e696. https://doi.org/10.1002/ctm2.696
You, Y., Zeng, Z., Zheng, J., Zhao, J., Luo, F., Chen, Y., Xie, M., Lui, X., y Wei, H. (2022). The toxicity response of Coccinella septempunctata L. (Coleoptera: Coccinellidae) after exposure to sublethal concentrations of acetamiprid. Agriculture, 12(10), 1642. https://doi.org/10.3390/agriculture12101642
Zelaya-Molina, L. X., Chávez-Díaz, I. F., de los Santos-Villalobos, S., Cruz-Cárdenas, C. I., Ruíz-Ramírez, S., y Rojas-Anaya, E. (2022). Control biológico de plagas en la agricultura mexicana. Revista Mexicana de Ciencias Agrícolas, 13(27), 69-79. https://doi.org/10.29312/remexca.v13i27.3251
Notas de autor
danielazambrano20@hotmail.com
Información adicional
e-location: e6021