Dossier: Suelo y microorganismos
Actividad promotora del crecimiento vegetal de cuatro hongos endófitos aislados desde Araucaria araucana
Plant growth-promoting activity of four endophytic fungi isolated from Araucaria araucana

Actividad promotora del crecimiento vegetal de cuatro hongos endófitos aislados desde Araucaria araucana
Siembra, vol. 11, núm. 2, e6802, 2024
Universidad Central del Ecuador

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Recepción: 15 Junio 2024
Revisado: 26 Julio 2024
Aprobación: 02 Octubre 2024
Financiamiento
Fuente: ANID/FONDECYT
Nº de contrato: 11220305
Resumen: Los hongos endófitos [HE] son microorganismos que viven dentro de las plantas sin causar daño aparente, y colaboran en la síntesis de sustancias reguladoras del crecimiento, fijación de nitrógeno, protección contra patógenos, entre otros beneficios para las plantas. En este estudio se determinó la capacidad de producción de ácido indol acético [AIA] y de solubilización de fosfato, en cuatro cepas de HE aislados desde raíces de Araucaria araucana y se evaluó su efecto sobre la promoción del crecimiento en Nicotiana tabacum, utilizada como planta modelo. Los hongos fueron identificados como Phialocephala fortinii (cepa E-1), Penicillium melinii (cepa E-13), Umbelopsis dimorpha (cepa E-14) y Preussia cymatomera (cepa E-36). El AIA fue determinado utilizando el método de Salkowski, usando caldo de papa dextrosa [CPD] suplementado con 10 mg L-1 de L-triptófano, como precursor para la síntesis de AIA, y la producción fue expresada en mg L-1. La solubilización de fosfato fue determinada en medio sólido Pikovskaya, evaluando los halos formados en el medio y calculando el índice de eficiencia relativa de solubilización [ERS]. Finalmente, la estimulación del crecimiento vegetal por los HE en un co-cultivo con las plántulas de N. tabacum, en condiciones in vitro, fue evaluada a través de la producción de biomasa aérea y radicular de las plántulas. Los mayores valores de AIA y ERS fueron obtenidos con U. dimorpha (52,29 y 3,36 mg L-1, respectivamente). Los hongos utilizados promovieron el crecimiento de la biomasa aérea y radicular de las plantas de N. tabacum en condiciones in vitro, obteniéndose la mayor producción de biomasa seca total (aérea y radicular) con U. dimorpha, con 188,5 mg, valor significativamente mayor (p < 0,01) que el de las plantas control (8,85 mg). A través de este estudio se infiere que los HE aislados de A. araucana promueven el crecimiento vegetal y brindan beneficios para las plantas mediante la producción de fitohormonas.
Palabras clave: crecimiento vegetal, Umbelopsis dimorpha, AIA, solubilización de fosfato.
Abstract: Endophytic fungi [EF] are microorganisms that reside within plant tissues without causing apparent damage. They play a role in synthesizing growth-regulating substances, nitrogen fixation, protection against pathogens, and other benefits for plants. In this study, we evaluated the capacity of Indole Acetic Acid [IAA] production and phosphate solubilization in four strains of EF isolated from roots of Araucaria Araucana, and assessed their effect on the growth promotion of Nicotiana tabacum, as a model plant. Fungi were identified as Phialocephala fortinii (strain E-1), Penicillium melinii (strain E-13), Umbelopsis dimorpha (strain E-14), and Preussia cymatomera (strain E-36). The concentration of IAA was determined using the Salkowski method using potato dextrose broth [PDB] supplemented with 10 mg L-1 L-tryptophan, as a precursor for IAA synthesis. The yield was expressed in mg L-1. The capacity of phosphate solubilization was determined in Pikovskaya solid medium through the evaluation of halos formed in the medium and calculating the relative solubilization efficiency ratio [RSE]. Finally, we evaluated the effect of EF in a co-culture with N. tabacum seedlings under in vitro conditions measuring aerial and root biomass of the seedlings. The highest values for IAA and RSE were observed in the case of U. dimorpha (52.29 and 3.36 mg L-1, respectively). All fungi used promoted the growth of both aerial and root biomass of N. tabacum plants under in vitro conditions, obtaining the highest production of total dry biomass (aerial and root) with U. dimorpha, with a value of 188.5 mg, which was significantly higher (p < 0,01) than that of control plants (8.85 mg). Our findings suggest that EF isolated from A. araucana has the potential to promote plant growth and provide benefits to plants through the production of phytohormones.
Keywords: plant growth, Umbelopsis dimorpha, IAA, phosphate solubilization.
1. Introducción
Los hongos promotores del crecimiento vegetal [PGPF, del inglés Plant growth-promoting fungi] son microorganismos residentes en el suelo de la rizósfera de plantas, y se les conoce por mejorar los mecanismos de defensa y el crecimiento de ellas (Parra Martínez, 2023). Los PGPF desempeñan diversas funciones en las plantas, como potenciales biocontroladores al competir por espacio y nutrientes, producción de hormonas de crecimiento, solubilización de minerales, resistencia micoparasítica y saprofítica, colonización de raíces y resistencia sistémica inducida [RSI] en las plantas (Adedayo y Babalola, 2023). Entre las especies promotoras del crecimiento vegetal, se encuentran los hongos endófitos [HE].
Los HE se definen como microorganismos que pasan la mayor parte o todo su ciclo de vida colonizando los tejidos de la planta hospedera, sin causar un daño evidente, incluso en algunos casos son necesarios para la sobrevivencia de las especies vegetales (Collinge et al., 2022; Ortega et al., 2020). Esta interacción entre los HE y su huésped es descrita como mutualista (Card et al., 2016), ya que los HE brindan beneficios a su huésped, lo que excluiría de esta definición a hongos patógenos y/o saprófitos. Así, microorganismos actualmente considerados como endófitos (bacterias, hongos, virus, protozoos), pero que en algún momento causan efectos nocivos en la planta huésped, deberían ser clasificados de otra manera (Le Cocq et al., 2017). Mientras la interacción hongo-planta se mantenga equilibrada, el hongo obtiene nutrientes de la planta hospedante y, a cambio, ofrece beneficios para ella (Bamisile et al., 2018). Entre estos beneficios se ha descrito que los HE pueden aportar a las plantas un mayor contenido de nutrientes, protección, estimulación del crecimiento radicular y aéreo, y aumento de la tolerancia al estrés (biótico y abiótico) al producir metabolitos secundarios con una amplia diversidad estructural que les brindan a las plantas protección y resistencia contra herbívoros, microorganismos patógenos y distintos estreses ambientales (Sánchez-Fernández et al., 2013). Estos microorganismos tienen la capacidad de sintetizar hormonas vegetales que son moléculas que controlan la mayoría de los procesos fisiológicos y bioquímicos, como la división celular, el crecimiento, la diferenciación de los órganos aéreos y de las raíces de las plantas (Morocho y Leiva-Mora, 2019). Una de las fitohormonas secretada por algunos HE es el ácido indol acético [AIA], hormona importante en los procesos relacionados con el crecimiento de las plantas, como la elongación celular, el desarrollo del tejido vascular y la dominancia apical (Andrade Ayala et al., 2020; Fu et al., 2015). Gran parte de los mecanismos y regulaciones del crecimiento y desarrollo de las plantas se produce gracias a las fitohormonas, siendo las principales las auxinas, citoquininas, giberelinas, ácido abscísico, etileno y ácido salicílico. Las auxinas incluyen un grupo de moléculas importantes que se encuentran en plantas, hongos y bacterias; el tipo de auxina más abundante en la naturaleza es el AIA. Otras, como el ácido indol propiónico [4-CI-IAA] y el ácido indol-3-butírico [AIB], se encuentran en menores concentraciones en las plantas (Báez-Pérez et al., 2015). Se han realizado estudios en hongos productores de AIA, como es el caso del género Trichoderma, donde fueron estudiadas 35 cepas (Trichoderma sp.) debido al potencial del hongo para sintetizar esta hormona (Ortuño et al., 2013).
Otro de los aspectos relevantes tiene que ver con la capacidad que tienen los HE para solubilizar nutrientes, mecanismo que pueden desarrollar por la liberación de ácidos orgánicos o por la secreción de enzimas (fosfatasas ácidas o alcalinas). Esto es fundamental cuando se sabe que el fósforo [P] es un macronutriente primario para el crecimiento de las plantas y cada vez más escaso, incluso se piensa que su uso como fertilizante comercial estaría acotado al presente siglo (Mogollón et al., 2018), siendo preponderante buscar alternativas para la solubilización de este nutriente. Al respecto, la condición en que se encuentra el P en los suelos de Chile, asociado a materia orgánica, precipitado con compuestos de hierro, aluminio o calcio, o inmovilizado, adsorbido en partículas de arcilla, hacen necesaria la búsqueda de alternativas ecológicamente aceptables para estos fines. Es aquí donde los HE pueden jugar un rol fundamental en el ciclaje del P en los suelos, ya que tienen la habilidad de transformar el fosfato orgánico e inorgánico al romper los enlaces que forma el P con los iones metálicos de hierro, calcio y aluminio y así transformarlo en formas solubles y disponibles (Restrepo-Franco et al., 2015).
Araucaria araucana, una especie endémica de los bosques templados de la zona centro-sur y sur de Chile y de la Patagonia argentina, se caracteriza por su longevidad y su gran importancia cultural y científica. Además de ser un valioso recurso económico para los pueblos originarios que viven de la recolección y uso de sus semillas, como fuente de alimentación y de ingresos, es un recurso genético único en el mundo (Hermann, 2006). El microbioma de A. araucana ha sido poco estudiado, lo que ha incrementado el interés por identificar las especies fúngicas que habitan en su entorno. Estas especies, que provienen de ambientes extremos y cambiantes, podrían ofrecer un gran potencial biotecnológico (Alarcón et al., 2020). En el estudio realizado por Chávez et al. (2023) se aislaron diferentes HE y micorrícicos arbusculares de A. araucana, los cuales se inocularon en plántulas de la misma especie sometidas a estrés hídrico. Los resultados mostraron una notable mejora en la resistencia al estrés hídrico de las plántulas inoculadas, así como cambios favorables en la morfología y fisiología de las plantas (Chávez et al., 2023). Por este motivo, resulta de interés evaluar si esta interacción planta-hongo es beneficiosa también para otras especies vegetales como las de interés agrícola.
En este contexto, la hipótesis planteada aquí sugiere que los HE aislados de las raíces de Araucaria araucana (especie vegetal milenaria), estimulan el crecimiento vegetal a través de la producción de AIA y solubilización de fosfato. El objetivo de este trabajo fue comprobar la capacidad promotora del crecimiento vegetal de cuatro cepas de HE mediante la producción de fitohormonas y solubilización de fosfato. Para esto, la investigación se centró en determinar la solubilización de fosfato en medio sólido y la concentración de AIA producido por los HE en fermentación líquida. Adicionalmente, se evaluó la estimulación del crecimiento en Nicotiana tabacum (como planta modelo) en co-cultivo con los HE, a través de la determinación de la biomasa de las plantas. Estos datos sentarán las bases para posteriores ensayos de optimización, donde algunas condiciones de cultivo (pH del medio, tiempo de cultivo, concentración de triptófano) serán probadas para aumentar la producción de AIA.
2. Materiales y Métodos
2.1. Aislamiento y activación de cepas
Los HE utilizados en este estudio fueron aislados desde las raíces de A. araucana (Chávez et al., 2023). Para ello, las raíces de las plantas se lavaron con agua de grifo, cuidadosamente, para eliminar los restos de tierra. Luego, las raíces se cortaron en trozos de 0,5 cm (aproximadamente) y se esterilizaron superficialmente con etanol al 70 % durante 1 min, seguido de hipoclorito de sodio al 10 % durante 10 min. Las raíces esterilizadas superficialmente fueron lavadas con agua destilada estéril, secadas y luego distribuidas en placas de Petri que contenían extracto de malta y agar [EMA] al 1 % m/v. Las muestras se incubaron a 24 °C durante 7 días, y a partir de ellas se aislaron y purificaron las cepas fúngicas, que luego fueron identificadas molecularmente como Phialocephala fortinii [cepa E-1], Penicilium melinii [cepa E-13], Umbelopsis dimorpha [cepa E-14] y Preussia cymatomera [cepa E-36], todas pertenecientes a la colección de cultivos del Laboratorio de Investigación de Micorrizas y Aplicaciones Biotecnológicas de Hongos [LIMAB], Universidad de Concepción, Campus Los Ángeles. Estas cepas fueron reactivadas en medio de cultivo EMA al 1 % m/v, colocando un disco de agar/micelio de 0,5 cm en el centro de las placas, que luego se incubaron a 24 ± 1 °C en completa oscuridad durante un periodo de 10 días.
2.2. Evaluación de ácido indol acético (AIA)
Para determinar la producción de auxinas por las diferentes cepas estudiadas se siguió la metodología descrita por Shahab et al. (2009). Se utilizó el medio de cultivo caldo de papa dextrosa (PD, Merck) suplementado con L-triptófano, 10 mg L-1. Matraces Erlenmeyer de 100 mL, que contenían 25 mL del medio de cultivo, se inocularon con 2 discos de agar micelio (5 mm) y se incubaron por 7 días en agitación a 120 rpm y 24 ± 1 °C. Los matraces se inocularon en triplicado por cada cepa fúngica, y se prepararon controles de los que contenían el medio de cultivo sin inocular. Mediante filtración al vacío se recuperó el caldo para determinar la concentración de auxinas del tipo AIA (Shahab et al., 2009). Para ello, se preparó el reactivo de Salkowski, mediante una solución de 15 mL de FeCl3 x 6H2O (0,5 M) a la que se añadió 300 mL de H2SO4 (98 % v/v) y 500 mL de agua destilada (Glickmann y Dessaux, 1995). Luego, 1 mL de sobrenadante se mezcló con el reactivo de Salkowski, dejando reposar a temperatura ambiente en condiciones de oscuridad por 30 min; posteriormente se midió la absorbancia a 530 nm en un espectrofotómetro (TU-1810 Split Beam UV-VIS). Para cuantificar la producción de auxinas se preparó una curva de calibración a partir de concentraciones conocidas de AIA (10-50 mg L-1).
2.3. Solubilización de fosfato medio de cultivo sólido
La solubilización de fosfato fue evaluada en medio de cultivo sólido para las cuatro cepas de HE mencionadas en 2.1. Se ocupó el medio de cultivo Pikovskaya [PVK] (Sanchez-Gonzalez et al., 2022) que contiene: (NH4)2SO4 (0,5 g), KCl (0,2 g), MgSO4 x 7H2O (0,1 g), MnSO4 · H2O (0,004 g), NaCl (0,2 g), D–Glucosa (10 g), FeSO4 x 7H2O (0,002 g), Ca3 (PO4)2 (5g), extracto de levadura (0,5 g), cloranfenicol (0,1 g) y agar 1 % m/v. El medio fue esterilizado a 121 °C por 15 min y, una vez tibio, 25 mL del medio fueron dispensados en placas Petri bajo cámara de flujo laminar. Luego de la solidificación del medio PVK se inocularon discos agar/micelio de 0,5 cm de diámetro en el centro de las placas Petri. Las placas se incubaron en oscuridad a 24 ± 1 °C y cada 5 días fueron medidos el diámetro (mm) de las colonias y el halo de solubilización alrededor de las colonias, que indica la solubilización del fosfato insoluble del medio de cultivo. Para estimar el fosfato solubilizado se aplicó el índice de eficiencia relativa de solubilización [ERS], calculado mediante la ecuación [1] descrita por Romero-Fernandez et al. (2018).
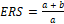 [1]
[1]donde:
-
a = diámetro de la colonia (mm)
-
b = diámetro del halo de solubilización (mm)
Para categorizar el nivel de solubilización de las cepas, se utilizó el criterio establecido por Silva-Filho y Vidor (2000), que indica que la capacidad de solubilización se considera baja cuando el ERS es inferior a 2, media si es superior a 2 e inferior a 3, y alta si es superior a 3.
2.4. Hongos endófitos como promotores de crecimiento vegetal
2.4.1. Material vegetal y condiciones de crecimiento
Para el co-cultivo se utilizaron semillas de Nicotiana tabacum y las cepas de hongos descritas anteriormente en el apartado 2.1. Las semillas se desinfectaron superficialmente mediante tratamiento con etanol al 70 % por 1 min, seguido de inmersión en hipoclorito de sodio 3 % y una gota de Tween 20 durante 10 min, se agitó en vortex cada 2 min. Finalmente, se procedió a enjuagar con agua destilada estéril por tres veces consecutivos, previo a la siembra. Para la germinación de las semillas se preparó un medio basal Murashige & Skoog [MS] completo, suplementado con sacarosa 3 % y agar 0,7 %, se ajustó el pH a 5,6 y se procedió a esterilizar durante 15 min a 1 atm de presión y 121 °C (Vilariño-Rodríguez, 2022).
2.4.2. Cocultivos de plantas y hongos
Luego de que las plantas de tabaco germinaron se transfirieron a una placa cuadrada de agar con medio MS suplementado con 0,5 % de sacarosa (p/v), momento en el cual se inocularon con los hongos. Dos discos de agar micelio (5 mm de diámetro) cortados con un sacabocados estéril, se colocaron asépticamente a una distancia de 1,5 cm entre ellos y a 5 cm de distancia de la radícula (3 plántulas por placa), según metodología de Dovana et al. (2015). La parte inferior de la placa fue cubierta con papel aluminio para evitar que la luz de la cámara de crecimiento afectara el micelio del hongo y la parte de la semilla germinada quedó expuesta a la luz. Se realizó un registro fotográfico cada 5 días posterior a la inoculación [dpi] para evaluar el crecimiento de las plantas. Al pasar 15 días se les realizó una cosecha destructiva, donde se evaluó crecimiento foliar y radicular para así determinar el peso seco (mg) de cada una de ellas.
2.5. Análisis estadísticos
La normalidad de los datos y la homogeneidad de la varianza se evaluaron mediante las pruebas de Shapiro-Wilk y Levene, respectivamente. La significación estadística se determinó usando test de Tukey (p < 0,05). Los análisis estadísticos se realizaron utilizando el software Statistica.v 10 (Statsoft). Cada ensayo se realizó por triplicado (n = 3).
3. Resultados y Discusión
3.1. Concentración de ácido indol acético [AIA]
Las cuatro cepas estudiadas tuvieron la capacidad de producir AIA y, como se puede observar en la Figura 1, la única cepa que presentó diferencias significativas en la producción de esta hormona fue la especie U. dimorpha cepa E-14 (52,29 mg L-1), y para las cepas E-1, E-13 y E-36 no se presentaron diferencias significativas entre ellas (17,2; 9,2 y 21,1 mg L-1, respectivamente). Al respecto, se han identificado seis vías biosintéticas diferentes de AIA, cinco de ellas dependientes de triptófano (Kejela, 2024). Por ende, los resultados sugieren que la producción de AIA por los distintos hongos estudiados tendría una ruta de síntesis dependiente de este precursor.
![Determinación de la producción de auxinas [AIA] de las diferentes cepas de HE evaluadas en medio PD. Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) y Preussia cymatomera (E-36). Los datos representan el promedio de tres réplicas ± la desviación estándar. Los valores seguidos por la misma letra no presentan diferencias significativas (p < 0,05).](https://revistadigital.uce.edu.ec/index.php/SIEMBRA/article/download/6802/version/6768/9178/41416/653877684016_gf6.png)
Figura 1
Determinación de la producción de auxinas [AIA] de las diferentes cepas de HE evaluadas en medio PD. Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) y Preussia cymatomera (E-36). Los datos representan el promedio de tres réplicas ± la desviación estándar. Los valores seguidos por la misma letra no presentan diferencias significativas (p < 0,05).
Figure 1. Determination of auxin production (IAA) for the different EF strains evaluated in PD medium. Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) and Preussia cymatomera (E-36). Data are expressed as mean ± standard deviation. Values followed by the same letter do not show significant differences (P < 0.05).
Se ha reportado que la cepa de U. dimorpha JSNL001-1 ha demostrado tener un efecto significativo en la mejora de la tasa de crecimiento y resistencia a la sequía en Dendrobium officinale (Xu, 2014). Este HE, además de producir AIA, como fue demostrado en este trabajo, tiene la capacidad de secretar distintos compuestos fenólicos y ácidos orgánicos que promueven el crecimiento vegetal, como fue descrito en el trabajo de Qin et al. (2018), donde se evaluó la producción de distintos metabolitos secretados por U. dimorpha en interacción con la planta Kadsura angustifolia, cambiando incluso algunos metabolitos propios de la planta huésped. Se ha estudiado que el aumento de raíces laterales puede aumentar la síntesis o producción de compuestos volátiles que activan genes relacionados a enzimas de la ruta biosintética de las auxinas (Hermosa et al., 2012). Los resultados muestran que los hongos estudiados poseen un potencial en la producción de AIA promoviendo el crecimiento vegetal de las raíces laterales y biomasa aérea (Figura 3). En Aspergillus awamori se encontró una producción de 24,2 μg mL-1 de AIA (Mehmood et al., 2019), y para el caso de Preussia sp. BSL-10, cultivado en medio de crecimiento Czapek durante 14 días, se detectó 1,64 ± 0,15 µg ml-1 de AIA (Al-Hosni et al., 2018). Además, en otros hongos (comestibles), como la especie Morchella importuna, se encontró una producción de AIA cercana a los 80 mg L-1 (Arroyo Marín, 2023), mayor a lo reportado en este estudio.
El microbioma de la rizósfera favorece el crecimiento de las plantas a través del establecimiento de interacciones mutualistas entre las plantas y los microorganismos, los cuales secretan diversos compuestos que promueven en la planta la síntesis de metabolitos beneficiosos para el desarrollo vegetal (Pascale et al., 2020). Sin embargo, los microorganismos también pueden, de manera independiente, sintetizar metabolitos que promueven el crecimiento de las plantas, tales como fitohormonas, sideróforos y enzimas que facilitan la solubilización de fosfato (Kumar, 2016). Naureen et al. (2022) aislaron 150 hongos desde la rizósfera y endósfera de la raíz de los pastos perennes Cymopogon jwarancusa y Panicum antidotale, y luego evaluaron la capacidad de los hongos para estimular el crecimiento vegetal de Arabidopsis thaliana (Col-0). Los autores demostraron que las cepas CJAN 1179 y PAAN 1135 de HE provocaron una significativa estimulación del crecimiento radicular, con la formación de raíces secundarias, después de 16 días de la inoculación de las plantas, y además que la cepa CJAN 1179 produjo 1.638 μg/mL de AIA en presencia de triptófano como precursor (Naureen et al., 2022).
3.2. Solubilización de fosfato
La evaluación de la actividad solubilizadora reveló una respuesta positiva con la formación de halos hialinos alrededor de las colonias (Figura 2b). El halo de solubilización se comenzó a presentar alrededor del tercer y quinto día, en el medio PVK. La cepa E-1 fue la única que no presentó halo de solubilización durante todo el ensayo (ESR = 1). Al respecto, Jones et al. (1991) y Whitelaw (1999) mencionan que la ausencia de halos de solubilización en los medios de cultivo sólidos no necesariamente indica que el organismo carezca de habilidad solubilizadora, sino que posiblemente este tipo de medios son insensibles para detectar la actividad de algunos microorganismos. Por lo cual es necesario recurrir a los medios líquidos para obtener resultados más precisos. Al respecto, Mikheev et al. (2022), utilizando P. fortinii, encontraron solubilización de fosfato atribuido a la producción de fitasa ácida (máximo de 6,91 ± 0,17 U el día 21 de cultivo en caldo PD) y capacidad para acumular polifosfatos en las células de las hifas, por lo que no se descarta que bajo otras condiciones de cultivo esta cepa (E-1) pudiese presentar actividad solubilizadora.
![a) Índice de eficiencia relativa de solubilización [ERS] a los 10 días de crecimiento. Los datos representan el promedio de tres réplicas ± la desviación estándar. Los valores seguidos por la misma letra no presentan diferencias significativas (p < 0,05). b) Halos de solubilización (zona hialina) de las cepas Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) y Preussia cymatomera (E-36), a los 10 días de crecimiento en medio PVK.](https://revistadigital.uce.edu.ec/index.php/SIEMBRA/article/download/6802/version/6768/9178/41417/653877684016_gf7.png)
Figura 2
a) Índice de eficiencia relativa de solubilización [ERS] a los 10 días de crecimiento. Los datos representan el promedio de tres réplicas ± la desviación estándar. Los valores seguidos por la misma letra no presentan diferencias significativas (p < 0,05). b) Halos de solubilización (zona hialina) de las cepas Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) y Preussia cymatomera (E-36), a los 10 días de crecimiento en medio PVK.
Figure 2. a) Relative solubilization efficiency ratio [RSE] at different growth periods (5, 10 and 15 days). Data are expressed as mean ± standard deviation. Values followed by the same letter do not show significant differences (P < 0.05). b) Solubilization halos (hyaline zone) of Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) and Preussia cymatomera (E-36), after 10 days of growth in PVK medium.
Se observó, a los diez días de crecimiento (Figura 2a), que E-14 fue la cepa con mayor índice de ERS (3,36), siendo significativamente mayor a los índices de solubilización presentados por las otras cepas. El desarrollo del halo alrededor de la colonia de los hongos se puede deber a la secreción de enzimas y/o ácidos orgánicos. Los ácidos orgánicos pueden acidificar las células del entorno circundante del hongo, provocando la liberación de iones P mediante la sustitución de H+ (unidos al fosfato), produciendo fósforo soluble para las plantas (Park et al., 2009; Saeed et al., 2021). Por otra parte, los ácidos orgánicos pueden quelar el catión unido al fosfato con sus grupos hidroxilos y carboxilos, haciendo que el P esté disponible en forma de ortofosfato (Picone y Zamuner, 2002). En un estudio donde se aislaron y caracterizaron hongos solubilizadores de fosfato para probarlos en una plantación de café, se analizaron 151 aislados, dentro de los cuales 21 tuvieron la capacidad de solubilizar fosfato, y dentro de ellos se encontraron especies de los géneros Umbelopsis y Penicillium, siendo las especies de Penicillium las que mostraron mayor capacidad de solubilización (Arias et al., 2023).
En relación con el índice de ERS los valores para las cepas E-13 y E-36 (2,23 y 2,18, respectivamente) fueron considerados medios y para la cepa E-14 (3,36) alto. Al respecto, los valores de este estudio son más altos que los señalados por Morales et al. (2011), quienes reportaron un índice de solubilización máximo de 1,3 para cepas de Penicillium albidum, P. thomii, P. restrictum, P. frequentans, Gliocladium roseum y Penicillium sp. Sin embargo, los valores obtenidos son menores que los reportados por Hajjam y Cherkaoui (2017), quienes han informado sobre valores de índices de solubilización de hasta 5,3 para Trichosporon beigelii. Arias Mota et al. (2022) informan de valores de ERS en el rango de 1,13-6,5 en hongos rizosféricos aislados de cafetales, lo que muestra la necesidad de realizar pruebas de selección iniciales para obtener cepas con potencial biotecnológico.
3.3. Estimulación del crecimiento vegetal
La biomasa aérea de las plantas de N. tabacum, inoculadas con los HE, mostró un aumento significativo respecto a las plantas control con todas las cepas estudiadas, destacando las cepas E-1 y E-14, que estimularon las mayores tasas de crecimiento, sin diferencias significativas entre ellas (39,8 y 37 mg, respectivamente). Los valores representaron un aumento de 5,78 y 6,21 veces respecto a las plantas control. A nivel radicular las cepas E-1, E-14 y E-36 mostraron diferencias significativas en relación con el control, obteniéndose el mayor peso seco con E-14 (72,5 ± 0,9 mg) (Figura 4). Esto podría estar relacionado con la mayor producción de AIA determinada con la cepa E-14 (Figura 1). Se conoce que las auxinas son responsables del comienzo de la formación de raíces laterales afectando el ciclo celular, activando células madre embrionarias (founder cells) presentes en el periciclo (Glick et al., 2012; Naureen et al., 2022), fenómeno que se vio evidenciado en las plantas de N. tabacum que desarrollaron raíces secundarias (Figura 3), desde los 10 dpi, a diferencia de las plantas control que no tuvieron desarrollo de raíces secundarias. En estudios similares donde se probó el hongo endófito Cyanodermella asteris en plantas de A. thaliana, las raíces presentaron un mayor crecimiento lateral, mayor biomasa y raíz principal más corta, en respuesta a las auxinas detectadas en las puntas de las raíces y primordios laterales (Jahn et al., 2021).

Figura 3
Crecimiento de N. tabacum a los 5, 10 y 15 días luego de la inoculación con las diferentes cepas de los HE: Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) y Preussia cymatomera (E-36).
Figure 3. Growth of N. tabacum at 5, 10 and 15 days after inoculation with different strains of EF Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) and Preussia cymatomera (E-36).

Figura 4
Biomasa aérea y radicular de las plantas de N. tabacum inoculadas con las diferentes cepas de los HE: Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) y Preussia cymatomera (E-36). Los datos representan el promedio de tres réplicas ± la desviación estándar. Los valores seguidos por la misma letra no presentan diferencias significativas (p < 0,05).
Figure 4. Aboveground and root biomass of N. tabacum plants inoculated with different strains of EF: Phialocephala fortinii (E-1), Penicillium melinii (E-13), Umbelopsis dimorpha (E-14) and Preussia cymatomera (E-36). Data are expressed as mean ± standard deviation. Values followed by the same letter do not show significant differences (p < 0.05).
La cepa E-13 no permitió el desarrollo radicular de las plantas debido a la abundante esporulación del hongo, que inhibió el crecimiento total de la planta (Figura 3), a pesar de esta cepa demostrar capacidad de secreción de AIA (Figura 1). En estudios realizados en P. citrinum aislado de Triticum aestivum L en condiciones de estrés por sequía (Kaur y Saxena, 2023), se determinó la producción de fitohormonas, como AIA, por este hongo. Los autores proponen que los hongos tolerantes a la sequía, como P. citrinum, activan los sistemas adaptativos del trigo y, a su vez, otorgan protección y estimulación del crecimiento vegetal (Kaur y Saxena, 2023). En otro estudio, donde se evaluó el hongo halotolerante P. olsonii en plantas de tabaco en cultivo hidropónico, se observó un incremento de la tolerancia a la salinidad, estimulando el crecimiento vegetal y reduciendo a la mitad el uso de fertilizantes químicos (Tarroum et al., 2022).
4. Conclusiones
Todas las especies/cepas evaluadas en este estudio presentaron producción de fitohormonas del tipo auxinas [AIA]. La especie Umbelopsis dimorpha (cepa E-14) fue la que presentó la mayor producción de AIA. Los HE tuvieron la capacidad de solubilizar fosfato, a excepción de Phialocephala fortinii (E-1), que no presentó capacidad de solubilización de fosfato en el medio PVK sólido. Los HE aislados de raíces de Araucaria araucana promovieron el crecimiento vegetal de N. tabacum mediante la producción de AIA y la solubilización de fosfato, lo que deja abierta la posibilidad de probar estos hongos o sus extractos fúngicos con otras plantas de interés agrícola y/o forestal.
Financiamiento
Esta investigación fue financiada por ANID/FONDECYT, número de subvención 11220305.
Contribuciones de los autores
-
Carolina Díaz-Valenzuela: conceptualización, investigación, metodología, software, análisis formal, redacción – borrador original, redacción - revisión y edición.
-
Christian Deramond Gálvez: investigación, metodología, curación de datos, análisis formal, software, redacción – borrador original, redacción - revisión y edición.
-
Ángela Machuca: recursos, supervisión, visualización, redacción - revisión y edición.
-
Daniel Eduardo Chávez Matamala: conceptualización, supervisión, validación, administración de proyectos, recursos, visualización, redacción – borrador original, redacción - revisión y edición.
Implicaciones éticas
Los autores declaran que las actividades de investigación se realizaron en el marco de las autorizaciones N° 02/2017 IX y N° 05/2022 IX.
Conflicto de interés
Los autores declaran que no existen conflictos de interés financieros o no financieros que podrían haber influido en el trabajo presentado en este artículo.
Agradecimientos
Agradecemos a la Administración de Parques Nacionales [CONAF] por las autorizaciones N° 02/2017 IX y N° 05/2022 IX, para realizar Actividades de Investigación en el Sistema Nacional de Áreas Silvestres Protegidas del Estado de Chile.
Referencias
Adedayo, A. A., y Babalola, O. O. (2023). Fungi that promote plant growth in the rhizosphere boost crop growth. Journal of Fungi, 9(2), 239. https://doi.org/10.3390/jof9020239
Alarcón, J., Márquez, S., Teunisse, G., Mendoza, C., Meneses, C., Baldini, A., Parra, P., Zamora, P., Boehmwald, F., y Castro-Nallar, E. (2020). Sequences of Endophytic fungal and bacterial communities from Araucaria araucana [(Molina) K. Koch, 1869] in the coastal and Andes Mountain Ranges, Chile. Microbiology Resource Announcements, 9(27). https://doi.org/10.1128/MRA.00544-20
Al-Hosni, K., Shahzad, R., Latif Khan, A., Muhammad Imran, Q., al Harrasi, A., al Rawahi, A., Asaf, S., Kang, S.-M., Yun, B.-W., y Lee, I.-J. (2018). Preussia sp. BSL-10 producing nitric oxide, gibberellins, and indole acetic acid and improving rice plant growth. Journal of Plant Interactions, 13(1), 112-118. https://doi.org/10.1080/17429145.2018.1432773
Andrade Ayala, M. del C. N., Hernandez Castillo, F. D., Laredo Alcala, E. I., Ledezma Pérez, A. S., Alvarado Canché, C. N., y Romero García, J. (2020). Efecto biológico de nanopartículas cargadas con ácido indolacético microbiano en parámetros morfométricos de tomate. Revista Mexicana de Ciencias Agrícolas, 11(3), 507-517. https://doi.org/10.29312/remexca.v11i3.1919
Arias Mota, R. M., Juárez González, A., Heredia Abarca, G., y de la Cruz Elizondo, Y. (2022). Capacidad fosfato solubilizadora de hongos rizosféricos provenientes de cafetales de Jilotepec, Veracruz. Alianzas y tendencias BUAP, 7(27), 69-86. https://doi.org/10.5281/zenodo.7094878
Arias, R. M., Heredia Abarca, G., del Carmen Perea Rojas, Y., de la Cruz Elizondo, Y., y García Guzman, K. Y. (2023). Selection and characterization of phosphate-solubilizing fungi and their effects on coffee plantations. Plants, 12(19), 3395. https://doi.org/10.3390/plants12193395
Arroyo Marín, M. J. (2023). Actividad promotora del crecimiento vegetal de especies de Morchella provenientes de bosque nativo y de plantaciones forestales del centro- sur de Chile. Universidad de Concepción. https://repositorio.udec.cl/handle/11594/10725
Báez-Pérez, A., González-Molina, L., Solís Moya, E., Bautista-Cruz, A., y Bernal-Alarcón, M. A. (2015). Efecto de la aplicación del ácido indol-3-butírico en la producción y calidad de trigo (Triticum aestivum L.). Revista Mexicana de Ciencias Agrícolas, 6(3), 523-537. http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-09342015000300007&lng=es&tlng=es.x
Bamisile, B. S., Dash, C. K., Akutse, K. S., Keppanan, R., y Wang, L. (2018). Fungal endophytes: beyond herbivore management. Frontiers in Microbiology, 9, 544. https://doi.org/10.3389/fmicb.2018.00544
Card, S., Johnson, L., Teasdale, S., y Caradus, J. (2016). Deciphering endophyte behaviour: the link between endophyte biology and efficacious biological control agents. FEMS Microbiology Ecology, 92(8), fiw114. https://doi.org/10.1093/femsec/fiw114
Chávez, D., Rivas, G., Machuca, Á., Santos, C., Deramond, C., Aroca, R., y Cornejo, P. (2023). Contribution of arbuscular mycorrhizal and endophytic fungi to drought tolerance in Araucaria araucana seedlings. Plants, 12(11), 2116. https://doi.org/10.3390/plants12112116
Collinge, D. B., Jensen, B., y Jørgensen, J. L. H. (2022). Fungal endophytes in plants and their relationship to plant disease. Current Opinion in Microbiology, 69, 102177. https://doi.org/10.1016/j.mib.2022.102177
Dovana, F., Mucciarelli, M., Mascarello, M., y Fusconi, A. (2015). In Vitro Morphogenesis of Arabidopsis to search for novel endophytic fungi modulating plant growth. PLOS ONE, 10(12), e0143353. https://doi.org/10.1371/journal.pone.0143353
Fu, S. F., Wei, J. Y., Chen, H. W., Liu, Y. Y., Lu, H. Y., y Chou, J. Y. (2015). Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signaling & Behavior, 10(8), e1048052. https://doi.org/10.1080/15592324.2015.1048052
Glick, B. R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica, 2012, 963401. https://doi.org/10.6064/2012/963401
Glickmann, E., y Dessaux, Y. (1995). A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Applied and environmental microbiology, 61(2), 793-796. https://doi.org/10.1128/aem.61.2.793-796.1995
Hajjam, Y. y Cherkaoui, S. (2017). The influence of phosphate solubilizing microorganisms on symbiotic nitrogen fixation: Perspectives for sustainable agriculture. Journal of Materials and Environmental Sciences, 8(3), 801-808. https://www.jmaterenvironsci.com/Journal/vol8-3.html
Hermann, T. M. (2006). Indigenous Knowledge and Management of Araucaria araucana forest in the Chilean Andes: Implications for Native Forest Conservation. Biodiversity and Conservation, 15, 647-662. https://doi.org/10.1007/s10531-005-2092-6
Hermosa, R., Viterbo, A., Chet, I., y Monte, E. (2012). Plant-beneficial effects of Trichoderma and of its genes. Microbiology, 158(1), 17-25. https://doi.org/10.1099/mic.0.052274-0
Jahn, L., Hofmann, U., y Ludwig-Müller, J. (2021). Indole-3-acetic acid is synthesized by the endophyte Cyanodermella asteris via a Tryptophan- dependent and independent way and mediates the interaction with a non-host plant. International Journal of Molecular Sciences, 22(5), 2651. https://doi.org/10.3390/ijms22052651
Jones, D., Smith, B. F. L., Wilson, M. J., y Goodman, B. A. (1991). Phosphate solubilizing fungi in a Scottish upland soil. Mycological Research, 95(9), 1090-1093. https://doi.org/10.1016/S0953-7562(09)80553-4
Kaur, R., y Saxena, S. (2023). Penicillium citrinum, a drought-tolerant endophytic fungus isolated from wheat (Triticum aestivum L.) leaves with plant growth-promoting abilities. Current microbiology, 80(5), 184. https://doi.org/10.1007/s00284-023-03283-3
Kejela, T. (2024). Phytohormone-producing Rhizobacteria and their role in plant growth. En B. Ali y J. Iqbal (eds.), New insights into phytohormones. IntechOpen. https://doi.org/10.5772/intechopen.1002823
Kumar, V. V. (2016). Plant growth-promoting microorganisms: interaction with Plants and Soil. En K. Hakeem, M. Akhtar, y S. Abdullah (eds.), Plant, Soil and Microbes (pp. 1-10). Springer. https://doi.org/10.1007/978-3-319-27455-3_1
Le Cocq, K., Gurr, S. J., Hirsch, P. R., y Mauchline, T. H. (2017). Exploitation of endophytes for sustainable agricultural intensification. Molecular Plant Pathology, 18(3), 469-473. https://doi.org/10.1111/mpp.12483
Mehmood, A., Hussain, A., Irshad, M., Hamayun, M., Iqbal, A., y Khan, N. (2019). In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis, 77, 225-235. https://doi.org/10.1007/s13199-018-0583-y
Mikheev, V. S., Struchkova, I. v., Ageyeva, M. N., Brilkina, A. A., y Berezina, E. v. (2022). The role of Phialocephala fortinii in improving plants’ phosphorus nutrition: New puzzle pieces. Journal of Fungi, 8(11), 1225. https://doi.org/10.3390/jof8111225
Mogollón, J. M., Beusen, A. H. W., van Grinsven, H. J. M., Westhoek, H., y Bouwman, A. F. (2018). Future agricultural phosphorus demand according to the shared socioeconomic pathways. Global Environmental Change, 50, 149-163. https://doi.org/10.1016/J.GLOENVCHA.2018.03.007
Morales, A., Alvear, M., Valenzuela, E., Castillo, C. E., y Borie, F. (2011). Screening, evaluation and selection of phosphate-solubilising fungi as potential biofertiliser. Journal of Soil Science and Plant Nutrition, 11(4), 89-103. https://dx.doi.org/10.4067/S0718-95162011000400007
Morocho, M. T., y Leiva-Mora, M. (2019). Microorganismos eficientes, propiedades funcionales y aplicaciones agrícolas. Centro Agrícola, 46(2), 93-103. http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0253-57852019000200093
Naureen, A., Nasim, F. H., Choudhary, M. S., Ashraf, M., Grundler, F. M. W., y Schleker, A. S. S. (2022). A new endophytic fungus CJAN1179 isolated from the Cholistan desert promotes lateral root growth in Arabidopsis and produces IAA through tryptophan-dependent pathway. Archives of Microbiology, 204(3), 181. https://doi.org/10.1007/s00203-022-02768-2
Ortega, H. E., Torres-Mendoza, D., y Cubilla-Ríos, L. (2020). Patents on endophytic fungi for agriculture and bio- and phytoremediation applications. Microorganisms, 8(8), 1237. https://doi.org/10.3390/microorganisms8081237
Ortuño, N, Miranda, C., y Claros, M. (2013). Selección de cepas de Trichoderma spp. generadoras de metabolitos secundarios de interés para su uso como promotor de crecimiento en plantas cultivadas. Journal of the Selva Andina Biosphere, 1(1), 16-24. https://sars.org.bo/index.php/jsab/article/view/134/173
Park, K.-H., Lee, C.-Y., y Son, H.-J. (2009). Mechanism of insoluble phosphate solubilization by Pseudomonas fluorescens RAF15 isolated from ginseng rhizosphere and its plant growth-promoting activities. Letters in Applied Microbiology, 49(2), 222–228. https://doi.org/10.1111/j.1472-765X.2009.02642.x
Parra Martínez, S. (2023). Microbiología del vermicompost. Universidad de los Andes. http://hdl.handle.net/1992/64905
Pascale, A., Proietti, S., Pantelides, I. S., y Stringlis, I. A. (2020). Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Frontiers in Plant Science, 10, 1741. https://doi.org/10.3389/fpls.2019.01741
Picone, L. I., y Zamuner, E. (2002). Fósforo orgánico y fertilidad fosfórica. Informaciones Agronómicas del Cono Sur, 16, 11-15. http://www.ipni.net/publication/ia-lacs.nsf/issue/IA-LACS-2002-4
Qin, D., Wang, L., Han, M., Wang, J., Song, H., Yan, X., Duan, X., y Dong, J. (2018). Effects of an endophytic fungus Umbelopsis dimorpha on the secondary metabolites of host-plant Kadsura angustifolia. Frontiers in Microbiology, 9, 2845. https://doi.org/10.3389/fmicb.2018.02845
Restrepo-Franco, G. M., Marulanda-Moreno, S., Fe-Pérez, Y. D. L., Díaz-de la Osa, A., Lucia-Baldani, V., y Hernández-Rodríguez, A. (2015). Bacterias solubilizadoras de fosfato y sus potencialidades de uso en la promoción del crecimiento de cultivos de importancia económica. Revista CENIC Ciencias Biológicas, 46(1), 63-76. https://revista.cnic.cu/index.php/RevBiol/article/view/95
Romero Fernández, A. de J., Arias Mota, R. M., y Mendoza Villarreal, R. (2018). Aislamiento y selección de hongos de suelo solubilizadores de fósforo nativos del estado de Coahuila, México. Acta Botánica Mexicana, (126), e1390. https://doi.org/10.21829/abm126.2019.1390
Saeed, Q., Xiukang, W., Haider, F. U., Kučerik, J., Mumtaz, M. Z., Holatko, J., Naseem, M., Kintl, A., Ejaz, M., Naveed, M., Brtnicky, M., y Mustafa, A. (2021). Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. International Journal of Molecular Sciences, 22(19), 10529. https://doi.org/10.3390/ijms221910529
Sánchez-Fernández, R. E., Sánchez-Ortiz, B. L., Sandoval-Espinosa, Y. K. M., Ulloa-Benítez, Á., Armendáriz-Guillén, B., García-Méndez, M. C., y Macías- Rubalcava, M. L. (2013). Hongos endófitos: fuente potencial de metabolitos secundarios bioactivos con utilidad en agricultura y medicina. TIP, Revista especializada en ciencias químico-biológicas, 16(2), 132-146. https://doi.org/10.1016/S1405-888X(13)72084-9
Sanchez-Gonzalez, Ma. E., Mora-Herrera, M. E., Wong-Villarreal, A., de La Portilla-López, N., Sanchez-Paz, L., Lugo, J., Vaca-Paulín, R., del Aguila, P., y Yañez-Ocampo, G. (2022). Effect of pH and carbon source on phosphate solubilization by bacterial strains in Pikovskaya medium. Microorganisms, 11(1), 49. https://doi.org/10.3390/microorganisms11010049
Shahab, S., Ahmed, N., y Khan, N. S. (2009). Indole acetic acid production and enhanced plant growth promotion by indigenous PSBs. African Journal of Agricultural Research, 4(11), 1312-1316. https://academicjournals.org/journal/AJAR/article-full-text-pdf/BCF994938188.pdf
Silva Filho, G. N., y Vidor, C. (2000). Solubilização de fostatos por microrganismos na presença de fontes de carbono. Revista Brasileira de Ciência Do Solo, 24(2), 311–319.
Tarroum, M., Romdhane, W. ben, Al-Qurainy, F., Ali, A. A. M., Al-Doss, A., Fki, L., y Hassairi, A. (2022). A novel PGPF Penicillium olsonii isolated from the rhizosphere of Aeluropus littoralis promotes plant growth, enhances salt stress tolerance, and reduces chemical fertilizers inputs in hydroponic system. Frontiers in Microbiology, 13, 996054. https://doi.org/10.3389/fmicb.2022.996054
Vilariño-Rodríguez, S. (2022). Avances en la producción de metabolitos secundarios de interés farmacológico a partir de material vegetal de Stevia rebaudiana, Bert. Universidad de Sevilla. https://idus.us.es/handle/11441/142948
Whitelaw, M. A. (1999). Growth Promotion of plants inoculated with phosphate-solubilizing fungi. Advances in Agronomy, 69, 99-151. https://doi.org/10.1016/S0065-2113(08)60948-7
Xu, C. (2014). Umbelopsis dimorpha and application of Umbelopsis dimorpha in promoting growth and improving drought resistance of Dendrobium officinale (Patente de China, N.° CN104195054A). https://patents.google.com/patent/CN104195054A/en
Notas de autor
danielchavez@udec.cl
Información adicional
e-location: e6802