Dossier: Suelo y microorganismos
Comunidades de hongos micorrízicos arbusculares (HMA) asociados al cultivo de mango cv. azúcar (Mangifera indica L.) en Magdalena, Colombia
Arbuscular mycorrhizal fungal (AMF) communities associated with mango cv. sugar (Mangifera indica L.) crops in Magdalena, Colombia
 Marlon José Yacomelo-Hernández myacomelo@agrosavia.co
Francisco Fabián Carrascal-Pérez fcarrascal@agrosavia.co
Wilmar Alexander Wilches-Ortiz wwilches@agrosavia.co
Luciano Ramírez luramirez@agrosavia.co
Diana Paola Serralde-Ordoñez dserralde@agrosavia.co
Andrea María Peñaranda-Rolon apenarandarolon@gmail.com
Margarita Ramírez- Gómez mmramirez@agrosavia.co
Marlon José Yacomelo-Hernández myacomelo@agrosavia.co
Francisco Fabián Carrascal-Pérez fcarrascal@agrosavia.co
Wilmar Alexander Wilches-Ortiz wwilches@agrosavia.co
Luciano Ramírez luramirez@agrosavia.co
Diana Paola Serralde-Ordoñez dserralde@agrosavia.co
Andrea María Peñaranda-Rolon apenarandarolon@gmail.com
Margarita Ramírez- Gómez mmramirez@agrosavia.co
Comunidades de hongos micorrízicos arbusculares (HMA) asociados al cultivo de mango cv. azúcar (Mangifera indica L.) en Magdalena, Colombia
Siembra, vol. 11, núm. 2, e7178, 2024
Universidad Central del Ecuador

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
Recepción: 10 Septiembre 2024
Revisado: 21 Septiembre 2024
Aprobación: 24 Octubre 2024
Resumen: Los hongos micorrízicos arbusculares son uno de los organismos más abundantes en la rizósfera y se pueden encontrar en más de 200.000 especies de plantas. A pesar de su gran distribución en ecosistemas, hasta el momento solo se han descrito 322 especies. El objetivo del presente estudio fue caracterizar la diversidad de las comunidades de hongos micorrízicos arbusculares presentes en dos áreas productoras de mango cv. azúcar en el departamento de Magdalena, Colombia. Para ello se llevó a cabo: (i) la identificación de las morfoespecies presentes, (ii) la comparación de las comunidades a través de índices de diversidad, (iii) la determinación de las propiedades químicas de los suelos, y (iv) la relación entre estas propiedades con la riqueza de especies, seleccionadas mediante análisis de componentes principales (ACP). Como resultado se encontró un total de 25 morfoespecies, representadas en seis familias y diez géneros, permitiendo identificar tres morfoespecies claves: Cetrasporapellucida, Glomussinuosum y Gigasporamargarita como las especies más frecuentes. Los mejores valores en los índices de diversidad se encontraron en Santa Marta, siendo la finca San Pablo la que obtuvo los valores más altos en los índices de Shannon y Simpson (1,63 y 0,78, respectivamente), mientras que la finca Las Palmeras fue la que obtuvo el mayor valor en el índice de uniformidad de Pielou con un valor de 0,97. Reportándose por primera vez la ocurrencia de especies de HMA asociadas a cultivos de mango cv. azúcar en las localidades de Santa Marta y Ciénaga.
Palabras clave: índice de diversidad, Mangifera indica, micorriza, abundancia de esporas, riqueza de especies, mango cv. azúcar.
Abstract: Arbuscular mycorrhizal fungi represent one of the most prevalent group organisms in the rhizosphere. They are present in more than 200,000 plant species. Despite their pervasive distribution in ecosystems, only 322 species have been formally described to date. The present study aimed to characterize the diversity of AMF communities in two mango-producing areas in the Magdalena Department, Colombia. To this end, the following steps were taken: (i) the identification of the morphospecies present, (ii) the comparison of communities through diversity indices, (iii) the determination of soil chemical properties, and (iv) the relationship between these properties with species richness, selected using principal component analysis (PCA). The study yielded 25 morphospecies, distributed across six families and ten genera. Of these, three key morphospecies Cetrasporapellucida, Glomus sinuosum and Gigaspora margarita were identified as the most frequent. The highest diversity values were observed in Santa Marta, with the San Pablo farm demonstrating the highest values for Shannon and Simpson indices (1,63 and 0,78, respectively), while the Las Palmeras farm exhibited the highest value for the Pielou uniformity index (0,97). This study reports, for the first time, the occurrence of AMF species associated with mango cv. sugar crops in the localities of Santa Marta and Ciénaga.
Keywords: diversity index, Mangifera indica, mycorrhizas, spore abundance, species richness, sugar mango.
1. Introducción
La mayoría de las plantas terrestres forman algún tipo de asociación simbiótica con hongos micorrízicos (van der Heijden et al., 2015), de los cuales un 1,5 % pertenecen a micorrizas ericoides, 2,0 % a ectomicorrizas [EcM], 10 % a micorrizas de tipo orquidoides y un 72 % se asocia a hongos micorrízicos arbusculares [HMA] (Brundrett y Tedersoo, 2018), siendo esta última la asociación más antigua y extendida en todo el planeta. Todos estos hongos juegan un papel clave en los ecosistemas terrestres, ya que regulan el ciclo de diversos nutrientes, influyen en la estructura del suelo y en la funcionalidad de los ecosistemas (van der Heijden et al., 2015). Los HMA son uno de los grupos de organismos más abundantes en la rizósfera, el cual se encuentra asociado a más de 200.000 especies de plantas. A pesar de su gran distribución en los ecosistemas, hasta el momento solo se han descrito, oficialmente, 317 especies (Goto y Jobim, 2024).
La riqueza y composición de las comunidades de HMA dependen tanto de la planta hospedera como de las condiciones ambientales; en el caso de las plantas cultivadas, además, del manejo agronómico, ya que su intensificación conduce a una reducción de la riqueza natural de los HMA (Öpik et al., 2006; Verbruggen et al., 2010; de Pontes et al., 2017). La identificación de especies nativas de HMA, es un factor clave para comprender como los cambios en la composición de las comunidades y en la diversidad pueden afectar los ecosistemas (Cofré et al., 2019).
El trópico suramericano es reconocido por su alta diversidad biológica y microbiológica; en este se destaca la amplia riqueza de los HMA que pueden encontrarse en diversos ecosistemas asociados con diferentes tipos de plantas (angiospermas y gimnospermas). Por ejemplo, los bosques tropicales son ampliamente colonizados por HMA y ectomicorrizas (Cofré et al., 2019), sin embargo, esta diversidad ha sido poco estudiada. En una búsqueda de publicaciones disponibles (110 artículos) sobre la diversidad de morfoespecies de HMA en Sur América, para Colombia solo se encontraron seis artículos publicados, donde se identificaron seis géneros y 20 especies de HMA (Cofré et al., 2019). De acuerdo con Peña-Venegas y Vasco-Palacios (2019), Colombia fue uno de los primeros países en Suramérica en estudiar a los HMA, pero hasta la fecha no se ha encontrado ningún artículo que haga referencia a la diversidad de HMA asociados al cultivo de mango cv. azúcar (Mangiferaindica).
El mango es una fruta tropical originaria de la India. Este país es el mayor productor y cuenta con el 22 % del área total de cultivos frutales, lo que comprende 2,3 millones de hectáreas y una producción de 24 millones de toneladas (FAOSTAT, 2021). En Colombia, el cultivo de mango, en 2022, tuvo un rendimiento de 10,31 t ha-1. Los principales productores de este cultivo son los departamentos de Tolima, Cundinamarca y Magdalena (Ministerio de Agricultura y Desarrollo Rural [MADR], 2020). En el mercado local se encuentra principalmente como fruta fresca y algunas variedades tradicionales son utilizadas por la agroindustria. Las principales variedades sembradas a escala nacional son el mango común, chancleto, costeño, azúcar, manzano, reina y Tommy Atkins, esta última destinada a las exportaciones (Aguilar Corrales et al., 2018). De estas variedades, la cv. azúcar se cultiva en 2.978 ha, donde se tiene un rendimiento promedio de 13,87 t ha-1 (Agronet, 2020).
A escala mundial se han realizado algunos estudios para conocer la diversidad de HMA asociados al cultivo de mango (Abdelhalim et al., 2014; Belay et al., 2014; Cortés Mercado et al., 1996; Govindan et al., 2020; Jiang et al., 2020; Mohandas, 2012; Mullath et al., 2019). Sin embargo, en Colombia no hay publicaciones disponibles sobre la diversidad de las comunidades de HMA asociadas a suelo rizosférico en mango cv. azúcar. En esta medida, el presente estudio tuvo como objetivo principal caracterizar la diversidad de las comunidades de HMA presentes en dos zonas productoras de mango cv. azúcar en el departamento de Magdalena en los municipios de Santa Marta y Ciénaga. Para ello, se realizó: (i) la identificación de las morfoespecies presentes, (ii) comparación de las comunidades mediante índices de diversidad, (iii) determinación de las propiedades químicas de los suelos, y (iv) la relación entre esas propiedades químicas con la riqueza de especies seleccionadas mediante un análisis de componentes principales [ACP].
2. Materiales y Métodos
2.1. Área de estudio
El estudio se realizó en los municipios de Santa Marta (11°14′10″N y 74°12′06″W, a 2 m s.n.m. de altitud, con un clima semiárido cálido) y Ciénaga (11°00′24″N y 74°14′48″W, a 3 m s.n.m. de altitud, con clima semiárido cálido) del departamento de Magdalena, Colombia. En cada municipio se escogieron seis fincas productoras de mango cv. azúcar (Mangifera indica L.) para un total de 12 puntos de muestreo (árboles de mango cv. azúcar) (Figura 1). Para el caso de Santa Marta fueron tomados los puntos La Madona [LMa], San Pablo [SP], Avilé [A], La Milagrosa [LMi], Rafael Barrios [RB] y Las Palmeras [LP]. Para Ciénaga se tomaron los puntos Edén [E], El Volcán [EV], Por Fin [PF], Miche Mary [MM], Las Margaritas [LMar] y El Triunfo [ET]. Se seleccionaron al azar tres árboles de mango cv. azúcar en cada punto de muestreo tomando una muestra compuesta de aproximadamente 1 kg de suelo rizosférico por cada uno, cada muestra fue colectada en un radio aproximado de 2 m alrededor de cada árbol, con el fin de colectar la mayor cantidad de raíces primarias activas. Por otra parte, se tomó de cada muestra compuesta de suelo una porción (500 g, aproximadamente) para analizar las propiedades químicas.
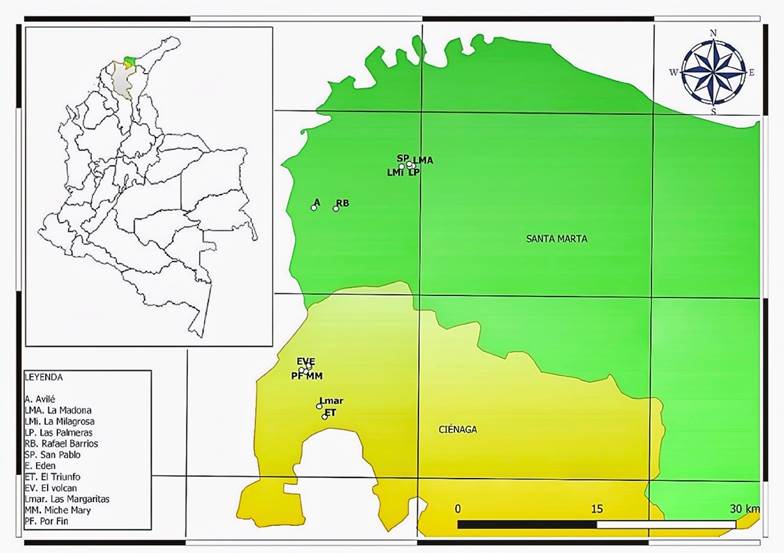
Figura 1
Fincas productoras de mango cv. azúcar en el departamento de Magdalena, Colombia, de donde fueron tomadas las muestras de suelo rizosférico para la obtención de esporas de HMA.
Figure 1. Mango cv. sugar producing farms in the department of Magdalena, Colombia, from which rhizospheric soils samples were taken to obtain AMF spores.
2.2. Análisis químico de suelo
Las muestras de suelo rizosférico fueron tomadas a una profundidad de 20 cm y fueron secadas a una temperatura de 40 °C, seguido de esto fueron molidas y tamizadas a 2 mm. Se midió el pH en suspensión suelo agua 1:2.5 (p/v); la capacidad de intercambio catiónico efectiva [CICE] por suma de cationes intercambiables; materia de orgánica [MO] por el método Walkley y Black; fósforo [P] disponible por el método de Bray II; K, Mg y Na intercambiables extraídos con CH3COONH4 1M pH 7,0; Cu, Fe, y Zn, disponibles extraídos por NTC5526:2007 extracción por Olsen modificado. Las muestras fueron analizadas en el laboratorio de química analítica del centro de investigación Tibaitatá de Agrosavia. Los análisis químicos se realizaron de las tres fincas donde se presentó la mayor abundancia de esporas de HMA de Santa Marta (LM, SP y A) y Ciénaga (E, EV y PF) (Tabla 1).
2.3. Aislamiento, cuantificación, caracterización morfológica, abundancia y riqueza de esporas de HMA
La caracterización morfológica de esporas y esporocarpos de HMA, a partir de muestras de suelo rizosférico de cultivos de mango cv. azúcar, inició con el aislamiento y cuantificación mediante decantado y suspensión de esporas en un gradiente de sacarosa, siguiendo la metodología propuesta por Gerdemann y Nicolson (1963).
Después del aislamiento y cuantificación, las esporas y esporocarpos fueron agrupados teniendo en cuenta características morfológicas como color, forma y tamaño. Se realizó el montaje para observación microscópica con ácido polivinil-láctico-glicerol [PVLG] (Koske y Tessier, 1983) mezclado con el reactivo de Melzer’s en proporción 1:1 (v / v) (Brundrett et al., 1994). Para el análisis microscópico de las características de las esporas se empleó un microscopio de luz (Zeiss; Primostar) con aumento de hasta 100X. La identificación se realizó con base en caracteres de la espora como: formación de tipo de espora (glomoide, acaulosporoide, entrophosporoide, gigasporoide y scutellosporoide), formación de esporocarpo, tamaño y color, estructura y tinción de paredes y características de germinación. Todas estas características morfológicas fueron contrastadas con la información disponible en los sitios web como el International Culture Collection of Arbuscular y Vesicular-Arbuscular Mycorrhizal Fungi (https://invam.ku.edu/), la de los profesores Janusz Blaszkowski (http://www.zor.zut.edu.pl/Glomeromycota_2/Home.html) y Bruno Tomio Goto (https://biologiademicorrizas.wixsite.com/glomeromycota/es), así como el sitio web de Schüßler (http://www.amf-phylogeny.com/), que ofrecen descripciones detalladas de gran parte de las especies de HMA aceptadas internacionalmente; así como publicaciones científicas que dan soporte para clasificación de géneros o especies (Chimal-Sánchez et al., 2020; Corazon-Guivin et al., 2019; de Souza et al., 2018; Lin et al., 2019; Oehl et al., 2008, 2011, 2019; Sieverding et al., 2014; Torres-Arias et al., 2017).
Después de la identificación a nivel morfológico de esporas y esporocarpos encontrados, se determinó la abundancia y riqueza de morfoespecies en cada muestra. La abundancia [A] se calculó como la cantidad de esporas de HMA 10 g-1 de suelo y la riqueza [R] como la cantidad de morfoespecies encontradas en cada muestra.
2.4. Estructura de las comunidades de HMA
La abundancia relativa [RD] se calculó mediante la relación porcentual entre especies de una muestra respecto al total de individuos encontrados. La frecuencia de ocurrencia de aislamientos [IF] se calculó determinando el porcentaje de muestras, de las cuales se aisló un género o una especie entre todas las muestras, reflejando su distribución, de acuerdo con la ecuación [1], propuesta por Brower et al. (1990):
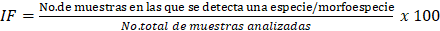 [1]
[1]Las morfoespecies se clasificaron como dominantes (IF > 50 %), muy comunes (31 < IF ≤ 50 %), comunes (10 < IF ≤ 30 %) y raras (IF < 10 %). Los índices de diversidad de Shanon-Wiener [H´], dominancia de Simpson [D]: y uniformidad de Pielou [J’] fueron calculados para cada muestra y entre las 12 muestras colectadas mediante las librerías vegan (Oksanen et al., 2020) y biodiversityR (Kindt y Coe, 2005) del software R® 4.3.2.
2.5. Análisis estadístico
Para los análisis químicos del suelo y densidad de esporas (en 10 g de suelo) se realizó un ANAVA y pruebas de comparación de medias de Tukey, posterior a la revisión de los supuestos de normalidad y homogeneidad de varianzas. Adicionalmente, se exploró la relación de las morfoespecies encontradas con las características químicas del suelo estandarizando las variables en un análisis de componentes principales [ACP] mediante software R ® 4.3.2 con las librerías factoextra, FactoMineR y ggplot2, en todos los casos se usó una p < 0,05.
3. Resultados
3.1. Análisis químico de suelos
Los puntos de muestreo presentaron valores de pH que van desde ligeramente ácido hasta ligeramente alcalino, con valores en el intervalo de 6,5 a 7,8. Los resultados obtenidos del análisis químico de suelos varió entre puntos de muestreos, encontrándose diferencias significativas entre ellos, siendo el punto A localizado en Santa Marta, el que presentó los valores más altos para la mayoría de elementos analizados, especialmente en los contenidos de fósforo, donde se registraron valores de 131,58 mg kg-1, valor muy superior al encontrado en los otros sitios de muestreo (Tabla 1). Sin embargo, este valor no influyó en la riqueza de HMA, pues los valores de menor riqueza fueron encontrados en los puntos de muestreo LP y RB (Tabla 2). Por otra parte, los resultados de la CE mostraron que estos suelos no presentaron condiciones de salinidad.
| Propiedades* | Unidad de medida | Sitios de muestreo | ||||||
| Santa Marta | Ciénaga | |||||||
| LM | SP | A | E | EV | PF | |||
| pH | 1:2.5 | 6,58 ± 0,14b | 6,47± 0,15b | 7,83 ± 0,15a | 7,67 ± 0,34a | 7,50 ± 0,21a | 7,11 ± 0,26ab | |
| CE | ds. m-1 | 0,26 ± 0,03abc | 0,24 ± 0,03bc | 0,70 ± 0,23a | 0,64 ± 0,18ab | 0,34 ± 0,08abc | 0,16 ± 0,01c | |
| MO | % | 1,13 ± 0,06ab | 1,26 ± 0,18ab | 1,43 ± 0,20a | 0,74 ± 0,17b | 1,03 ± 0,05ab | 0,81 ± 0,26ab | |
| Fósforo | mg. kg-1 | 10,29 ± 2,78b | 32,90 ± 9,17b | 131,58 ± 34,55a | 38,18 ± 14,19b | 46,04 ± 13,76b | 24,23 ± 10,74b | |
| Azufre | mg. kg-1 | 1,62± 0,19b | 1,69 ± 0,43ab | 3,13 ± 5,83a | 1,87 ± 1,61ab | 2,19 ± 0,81ab | 1,64 ± 0,48b | |
| CICE | cmol(+). kg-1 | 11,67± 0,43b | 10,12 ± 0,74b | 18,90 ± 2,87a | 11,75 ± 2,98b | 9,52 ± 0,78b | 14,67 ± 1,34ab | |
| Boro | mg. kg-1 | 0,07± 0,02b | 0,10 ± 0,03b | 0,90 ± 0,09a | 0,53 ± 0,23ab | 0,47 ± 0,24ab | 0,24 ± 0,08b | |
| Calcio | cmol(+). kg-1 | 9,14 ± 0,36ab | 7,97 ± 0,62b | 13,47 ± 2,06a | 7,86 ± 2,41b | 7,49 ± 0,82b | 11,37 ± 0,85ab | |
| Magnesio | cmol(+). kg-1 | 2,23 ± 0,11b | 1,76 ± 0,11b | 4,00 ± 0,79a | 2,52 ± 0,74ab | 1,28 ± 0,07b | 2,48 ± 0,38ab | |
| Potasio | cmol(+). kg-1 | 0,17 ± 0,01b | 0,19 ± 0,03b | 0,98 ± 0,10a | 0,40 ± 0,18b | 0,52 ± 0,28ab | 0,50 ± 0,12ab | |
| Sodio | cmol(+). kg-1 | 0,39 ± 0,01b | 0,43 ± 0,04a | 0,64 ±0,20ab | 0,96 ± 0,44a | 0,49 ± 0,06b | 0,57 ± 0,09ab | |
| Hierro | mg. kg-1 | 6,37± 10,91ab | 8,41 ± 19,60a | 2,79 ± 1,82c | 2,83 ± 1,05c | 3,19 ± 5,24c | 3,63 ± 3,05bc | |
| Cobre | mg. kg-1 | 1,00 ± 0,0a | 1,15 ± 0,20a | 1,29 ± 0,45a | 1,27 ± 0,86a | 2,12 ± 10,32a | 1,00 ± 0,0a | |
| Manganeso | mg. kg-1 | 1,72 ± 0,32a | 1,85 ± 0,43a | 1,11 ± 0,08a | 1,17 ± 0,11a | 1,09 ± 0,07a | 1,58 ± 0,42a | |
| Zinc | mg. kg-1 | 1,00 ± 0,0a | 1,39 ± 0,46a | 1,29 ± 0,64a | 1,59 ± 3,06a | 11,05 ± 13,76a | 1,00 ± 0,0a | |
Table 1. Chemical analysis of soils associated with mango cv. sugar (Mangifera indica) crops at the municipalities of Santa Marta and Ciénaga, Department of Magdalena, Colombia.
* CE: Conductividad eléctrica / Electrical Conductivity; MO: Materia orgánica / Organic Matter; CICE: Capacidad de intercambio catiónico / Cation Exchange Capacity. Letras diferentes significan diferencias significativas de acuerdo con la prueba de comparación de rangos múltiples de Tukey (p < 0,05) / Different letters mean significant differences according to Tukey's multiple range comparison test (p < 0.05).
| Familia / morfoespecie HMA | A | RD % | IF %* |
| Acaulosporaceae | |||
| Acaulospora excavata Ingleby y C. Walker | 10 | 1,5 | 25C |
| Acaulospora scrobiculata Trappe | 26 | 4 | 16,7C |
| Acaulospora mellea Spain y N.C. Schenck | 3 | 0,5 | 8,3R |
| Acaulospora sp1Gerdemann y Trappe | 22 | 3,4 | 8,3R |
| Ambisporaceae | |||
| Ambispora spWalker, Vestberg y Schuessler | 10 | 1,5 | 8,3R |
| Gigasporaceae | |||
| Gigaspora margarita W.N.Becker y I.R.Hall | 106 | 16,3 | 58,3D |
| Gigaspora sp1Gerd. y Trappe emend. C. Walker y F.E. Sanders | 5 | 0,8 | 16,7C |
| Glomeraceae | |||
| Funneliformis mosseae C. Walker y A. Schüssler | 4 | 0,6 | 8,3R |
| Funneliformis sp1Walker y Schüssler, emend. Oehl, Silva y Sieverding (Glomus group Aa1) | 2 | 0,3 | 8,3R |
| Glomus sinuosum R.T. Almeida y N.C. Schenck | 109 | 16,7 | 66,7D |
| Glomus taiwanense R.T. Almeida y N.C. Schenck | 30 | 4,6 | 25C |
| Glomus microcarpum Tul. y C. Tul | 58 | 8,9 | 16,7C |
| Glomus macrocarpum Tul. y C. Tul | 7 | 1,1 | 8,3R |
| Glomus sp1Tul. y C. Tul | 6 | 0,9 | 8,3R |
| Glomus sp2 | 14 | 2,1 | 33,3MC |
| Glomus sp3 | 8 | 1,2 | 8,3R |
| Glomus sp4 | 26 | 4 | 8,3R |
| Glomus sp5 | 1 | 0,2 | 8,3R |
| Glomus sp6 | 2 | 0,3 | 8,3R |
| Rhizoglomus fasciculatum G.A. Silva y Oehl | 3 | 0,5 | 8,3R |
| Rhizoglomus intraradices Sieverd., G.A. Silva y Oehl | 2 | 0,3 | 8,3R |
| Septoglomus constrictum G.A. Silva y Oehl | 40 | 6,1 | 8,3R |
| Racocetraceae | |||
| Cetraspora pellucida Oehl, F.A. Souza y Sieverd | 142 | 21,8 | 66,7D |
| Racocetra spOehl, F.A. Souza y Sieverding | 10 | 1,5 | 8,3R |
| Scutellosporacea | |||
| Scutellospora calospora C. Walker y F.E. Sanders | 6 | 0,9 | 16,7C |
| Total HFMA: | 652 | 100% | |
| 25 morfoespecies |
Table 2. Spore abundance, relative spore density, and i.solation frequency of AMFs identified from 12 sampling points of mango cv. sugar crops at two localities in Magdalena, Colombia.
* A: abundancia de esporas (10 g-1 de suelo) / spore abundance (10 g-1 soil); RD: densidad relativa / relative density; IF: frecuencia de ocurrencia de aislamientos / Isolate Frequency of Occurrence. Clase de especie según la IF (D: Dominante, MC: Muy común, C: Común y R: Raras) / Species class according to FI (D: Dominant, MC: Very common, C: Common and R: Rare).3.2. Caracterización morfológica, abundancia y riqueza de esporas de HMA
Un total de 652 esporas de HMA 10 g-1 de suelo fueron identificadas a partir de 36 muestras de los municipios de Santa Marta y Ciénaga, encontrándose 25 morfoespecies representadas en seis familias y 10 géneros (Tabla 2). La mayor riqueza de especies se registró en el punto de muestreo SP y la mayor abundancia de esporas (en 10 g-1 de suelo) en el A (Figura 2). La más alta abundancia de esporas se obtuvo de Cetraspora pellucida (Oehl, F.A. Souza & Sieverd.) con 142 esporas 10 g-1 de suelo, seguido de Glomus sinuosum (R.T. Almeida & N.C. Schenck), Gigaspora margarita (W.N. Becker & I.R. Hall), Glomus microcarpum (Tul. & C. Tul) y Septoglomus constrictum (G.A. Silva & Oehl) con 109, 106, 58 y 40 esporas 10 g-1 de suelo seco, respectivamente.

Figura 2
Riqueza de morfoespecies encontradas en cada punto de muestreo (A) y abundancia de esporas de HMA 10 g-1 de suelo (B)
Figure 2. Morphospecies richness registered at each sampling point (A) and AMF spore abundance in 10 g-1 soil (B)
En la Figura 3 se presentan algunas morfoespecies representativas aisladas de suelo rizosférico del cultivo de mango cv. azúcar en los municipios de Santa Marta y Ciénaga.

Figura 3
Esporas de HMA encontrados en suelos rizosféricos asociados al cultivo de mango cv. azúcar en los municipios de Santa Marta y Ciénaga. A. Acaulospora scrobiculata (Trappe); B. Acaulospora excavata (Ingleby & C. Walker); C. Cetraspora pellucida (Oehl, F.A. Souza & Sieverd.); D. Gigaspora margarita (W.N. Becker & I.R. Hall); E. Glomus microcarpum (Tul. & C. Tul); F. Glomus sinuosum (R.T. Almeida & N.C. Schenck); G. Glomus taiwanense (R.T. Almeida & N.C. Schenck); H. Rhizoglomus intraradices (Sieverd., G.A. Silva & Oehl); I. Rhizoglomus fasciculatum (G.A. Silva & Oehl); J. Septoglomus constrictum (G.A. Silva & Oehl).
Figure 3. AMF spores found in rhizospheric soils associated at mango cv. sugar (Mangifera indica) crops at the municipalities of Santa Marta and Ciénaga. A. Acaulospora scrobiculata (Trappe); B. Acaulospora excavata (Ingleby & C. Walker); C. Cetraspora pellucida (Oehl, F.A. Souza & Sieverd.); D. Gigaspora margarita (W.N.Becker & I.R.Hall); E. Glomus microcarpum (Tul. & C. Tul); F. Glomus sinuosum (R.T. Almeida & N.C. Schenck); G. Glomus taiwanense (R.T. Almeida & N.C. Schenck); H. Rhizoglomus intraradices (Sieverd., G.A. Silva & Oehl); I. Rhizoglomus fasciculatum (G.A. Silva & Oehl); J. Septoglomus constrictum (G.A. Silva & Oehl).
3.3. Estructura de la comunidad de HMA
Del total de familias, un 11 % pertenece a Acaulosporaceae, 2 % a Ambisporaceae, 21 % a Gigasporaceae, 45 % a Glomeraceae, 20 % a Racocetraceae y 1 % a Scutellosporaceae, siendo la familia Glomeraceae y Gigasporaceae las de mayor representación.
Se identificaron 14 morfoespecies a nivel de especie y 11 a nivel de género. El mayor número de morfoespecies encontradas pertenecen a los géneros Glomus (10) y Acaulospora (4) representando el 40 % y 16 % del total, respectivamente.
El 60 % de las morfoespecies de HMA encontradas se clasificaron como raras, 24 % como comunes y 4 % como muy comunes. Las morfoespecies dominantes presentes en 8 de los 12 puntos de muestreo estuvieron representadas por Cetraspora pellucida Oehl, F.A. Souza & Sieverd (IF = 66,7 %, RD = 21,8 %), Glomus sinuosum R.T. Almeida & N.C. Schenck (IF = 66,7 %, RD = 16,7 %) y Gigaspora margarita W.N. Becker & I.R. Hall (IF = 58,3 %, RD = 16,3 %) (Tabla 2).
Morfoespecies como Acaulospora melleaSpain & N.C. Schenck, Ambispora sp., F. mosseae C. Walker & A. Schüssler, G. microcarpum Tul & C. Tul, G. macrocarpum Tul & C. Tul, Glomus sp1., Racocetra sp. y Scutellospora calospora C. Walker & F.E. Sanders se encontraron solamente en Santa Marta, mientras que Acaulospora sp. 1., Gigaspora sp. 1., Funneliformis sp. 1., Glomus sp. 3, 4, 5 y 6, R. fasciculatum Sieverd.,G.A. Silva & Oehl y Rhizoglomus intraradices Sieverd., G.A. Silva & Oehl se encontraron únicamente en Ciénaga (Figura 4).

Figura 4
Abundancia relativa de morfoespecies de HMA en diferentes puntos de muestreo de Santa Marta y Ciénaga.
Figure 4. Relative abundance of AMF morphospecies at each sampling site in Santa Marta and Ciénaga.
La abundancia relativa en las dos localidades se caracterizó por estar en una mayor proporción representada por Cetrasporapellucida Oehl, F.A. Souza & Sieverd. en Santa Marta y por Glomus sinuosum R.T. Almeida & N.C. Schenck y Gigasporamargarita W.N. Becker & I.R. Hall en Ciénaga (Figura 4).
El mayor índice de diversidad de Shannon se presentó en el punto SP de Santa Marta con un valor de 1,96, seguido de LMar de Ciénaga con valor de 1,63, lo que indica buenas condiciones de conservación de los HMA encontrados. Contrario a esto, la menor diversidad de especies se encontró en el punto RB de Santa Marta con un valor del índice de Shannon de 0,59. La misma tendencia se observó para la dominancia con base en el índice de Simpson, en donde los puntos de muestreo SP y LMar fueron los que presentaron los mayores valores (0,83 y 0,78, respectivamente) y RB presentó los menores valores con una dominancia de 0,40 (Figura 5).
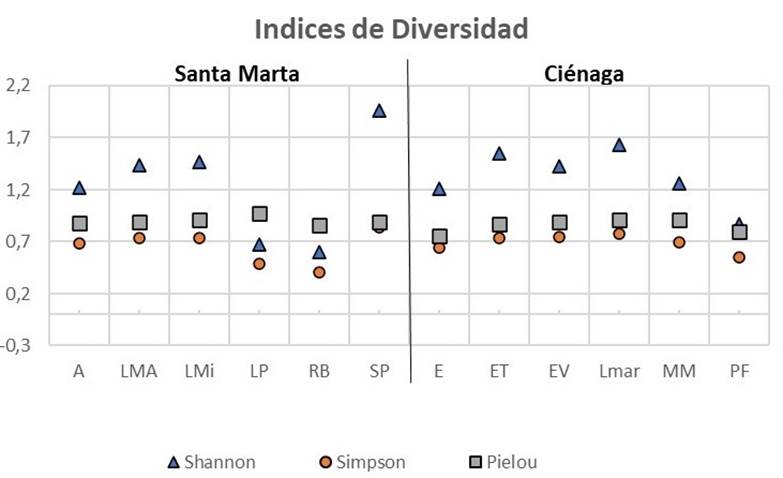
Figura 5
Índices de diversidad utilizados para describir las comunidades de HMA colectadas de diferentes puntos de muestreo en el cultivo de mango cv. azúcar en Magdalena, Colombia.
Figure 5. Diversity indices applied to describe AMF communities collected at different sampling points of mango cv. sugar (Mangifera indica) crops at Magdalena, Colombia.
El índice de uniformidad de especies de Pielou mide la proporción de la diversidad observada con relación a la máxima diversidad esperada. Este índice puede tener valores entre 0 y 1, donde 1 corresponde a situaciones donde todas las especies son igualmente abundantes (Magurran, 1988). El mayor índice de Pielou se presentó en LP de Santa Marta con un valor de 0,97, seguido por LMar con un valor de 0,90. Todas las morfoespecies de LP son las más proporcionadas en abundancia respecto a las demás fincas puesto que su valor Pielou fue el más cercano a 1 (Figura 5).
Por otra parte, los resultados del ACP mostraron la relación de las comunidades de HMA de los sitios estudiados con las características químicas de los suelos. Este análisis cuyos ejes principales explican el 63,1 % de la variabilidad, 39,8 % de la varianza total en el primer componente y 23,3 % en el segundo componente (Figura 6), se observó que Cetraspora pellucida Oehl, F.A. Souza & Sieverd, presentó una asociación positiva en orden de significancia con P (0,94), MO (0,72) y pH (0,57) (Figura 6). Por otra parte, Glomus sinuosum R.T. Almeida & N.C. Schenck presentó una correlación positiva en orden de significancia con S (0,98), P (0,97), B (0,95), K (0,94) pH (0,80), Mg (0,71), y Ca (0,62); mientras que, Gigaspora margarita W.N. Becker & I.R. Hall presentó una correlación negativa significativa con P (-0,75), B (-0,75), K (-0,70), S (-0,72) y pH (-0,64) (Figura 6).

Figura 6
Análisis de componentes principales con las morfoespecies encontradas en suelo rizosférico del cultivo de mango cv. azúcar y los atributos químicos del suelo en los diferentes puntos de muestreo. Ambispora sp. (Ambis sp), Glomus macrocarpum (Gl. macro), Funneliformis mosseae (Fun. moss) C. Walker & A. Schüssler, Acaulospora excavata (Acau. exc), Acaulospora mellea (Acau. mell), Glomus sp.1. (Gl. sp.1), Glomus sp.2. (Gl. sp.2), Glomus sp.5. (Gl. sp.5), Glomus sp.6. (Gl. sp.6). Cetraspora pellucida (Cetr. pell), Septoglomus constrictum (Septo. constr), Glomus sinuosum (Gl. sinuo), Rhizoglomus intaradices (Rhiz. intra), Rhizoglomus fasciculatum (Rhiz. fasc), Scutellospora calospora (Scut.calos), Glomus microcarpum (Gl. micro), Glomus taiwanensis (Gl. taiw), Gigaspora margarita (Gig. marg).
Figure 6. Principal component analysis with the morphospecies found in the rhizospheric soil of mango cv. sugar crop and soil chemical attributes at the different sampling points. Ambispora sp. (Ambis sp), Glomus macrocarpum (Gl.macro), Funneliformis mosseae (Fun.moss) C. Walker & A. Schüssler, Acaulospora excavata (Acau.exc), Acaulospora mellea (Acau.mell), Glomus sp1. (Gl.sp1), Glomus sp2. (Gl.sp2), Glomus sp5. (Gl.sp5), Glomus sp6. (Gl.sp6). Cetraspora pellucida (Cetr.pell), Septoglomus constrictum (Septo.constr), Glomus sinuosum (Gl.sinuo), Rhizoglomus intaradices (Rhiz.intra), Rhizoglomus fasciculatum (Rhiz.fasc), Scutellospora calospora (Scut.calos), Glomus microcarpum (Gl.micro), Glomus taiwanensis (Gl.taiw), Gigaspora margarita (Gig.marg).
4. Discusión
La diversidad de HMA presente en el cultivo de mango ha sido escasamente estudiada a escala mundial con pocos reportes en Colombia (Abdelhalim et al., 2014; Govindan et al., 2020; Jiang et al., 2020; Mullath et al., 2019). Este trabajo constituye el primer estudio para conocer la diversidad de HMA asociados al cultivo de mango cv. azúcar.
Teniendo en cuenta otros estudios realizados en mango comparados con los resultados obtenidos en este trabajo, la riqueza de morfoespecies puede considerarse alta, ya que se reportaron un total de 25 morfoespecies distribuidas en 6 familias y 10 géneros, en comparación a las encontradas en países como México, Sudán, Emiratos Árabes Unidos, India y Etiopía, en los que reportan entre 4 y 18 morfoespecies de HMA (Abdelhalim et al., 2014; Belay et al., 2014; Govindan et al., 2020; Mullath et al., 2019). Belay et al. (2014) y Abdelhalim et al. (2014) encontraron 18 morfoespecies en donde, de acuerdo con Abdelhalim et al. (2014), estuvieron distribuidas en una para Acaulospora, Ambispora, Archaeospora, Diversispora, Entrophospora y Septoglomus, tres para Claroideoglomus, dos para Funneliformis, cinco para para Glomus y dos para Paraglomus diferenciándose de Belay et al. (2014) en una morfoespecie para Rhizophagus, dos morfoespecies para Acaulospora, Claroideoglomus, cinco para Funneliformis. Por otra parte, Govindan et al. (2020) evaluaron el estatus de los HMA presentes en cinco variedades mejoradas y dos híbridos de mango, encontrando un total de diez especies pertenecientes a cinco géneros, una de Acaulospora y Gigaspora, dos de Claroideoglomus y Sclerocystis y cuatro de Glomus. Todos estos resultados sugieren que en suelos asociados al cultivo de mango cv. azúcar en el caribe colombiano, específicamente en las zonas de mayor producción (Santa Marta y Ciénaga), la diversidad de esporas de HMA es considerablemente superior, teniendo en cuenta que se lograron identificar 25 morfoespecies, distribuidas en una para Ambispora, Cetraspora, Racocetra, Septoglomus y Scutellospora, dos para Gigaspora, Rhizoglomus y Funneliformis, cuatro para Acaulospora y diez para Glomus.
En un estudio realizado en mango sobre dos sistemas de manejo, uno con manejo orgánico y otro con manejo convencional, se encontró un mayor número de morfoespecies en suelos con manejo orgánico, con un total de 6 especies representadas en 4 familias y 3 géneros, mientras que el manejo convencional reportó 4 especies distribuidas en 2 familias y 2 géneros (Mullath et al., 2019). Esta situación muestra que el manejo de los cultivos puede afectar las comunidades de HMA.
Otro factor que puede influenciar el comportamiento de las comunidades de HMA presentes es la etapa vegetativa de la planta y el genotipo cultivado. Estudios realizados por Adriano Anaya et al. (2008) en 4 materiales de mango observaron que la densidad de esporas puede variar con la etapa vegetativa y el material vegetal cultivado. La densidad de esporas encontrada en este estudio fue entre baja y media, con valores que oscilaron entre 17 y 126 esporas 10 g-1de suelo (Adriano Anaya et al., 2008). A diferencia de los estudios realizados por Cortés Mercado et al. (1996) en México y por Jiang et al. (2020) en China con valores encontrados de hasta 147,3 y 103 esporas g-1 de suelo seco, respectivamente.
Se ha demostrado que en los géneros pertenecientes al orden Gigasporales, gran parte de su estrategia de multiplicación es a partir de esporas y esta es una razón por la cual no pueden afrontar los ecosistemas perturbados (Abdelhalim et al., 2014). Por lo tanto, en este estudio las morfoespecies Gigasporamargarita y Cetraspora pellucida, pertenecientes al orden Gigasporales fueron las más frecuentes, debido a que en los suelos donde se cultiva mango cv. azúcar (Mangiferaindica) en el departamento de Magdalena son de textura arenosa y con muy poca perturbación ambiental. Estudios realizados han demostrado que la densidad de esporas de las especies de Gigaspora y Scutellospora (algunos reclasificados como Cetraspora) se reducen sustancialmente con labranza convencional ya que sus esporas son de mayor tamaño que las de otros órdenes y familias de HMA y estas pueden ser más susceptibles al daño físico (Allen et al., 2003; Boddington y Dodd, 2000; Lekberg et al., 2007). De La Providencia et al. (2005) encontraron abundantes esporas del orden Gigasporales en suelos arenosos, pero no en vertisoles arcillosos en donde, por los repetidos procesos de expansión y contracción de estos suelos, podrían afectar drásticamente las hifas de los Gigasporales.
Al igual que en este estudio, en suelos de la India también se ha encontrado a Gigaspora margarita como una de las morfoespecies más frecuentes (IF = 57,1 %) (Govindan et al., 2020). Adicionalmente, esta morfoespecie ha sido evaluada en injertos de mango bajo condiciones de invernadero con otras especies de HMA y se ha encontrado que Gi. margarita fue la que mejor respuesta ha generado en cuanto a crecimiento (altura de la planta, diámetro del tallo y biomasa) y nutrición (absorción de fósforo) de la planta, así como la reducción de mortalidad del injerto en condiciones de invernadero para ser llevado a campo (Reddy y Bagyaraj, 1994).
A pesar de las condiciones de los suelos en donde se cultiva mango cv. azúcar en el departamento de Magdalena, se observó que se presenta en forma natural la asociación simbiótica entre micorrizas – plantas de mango, encontrándose a Gigasporamargarita W.N. Becker & I.R. Hall, Glomussinuosum R.T. Almeida & N.C. Schenck y Cetraspora pellucida Oehl, F.A. Souza & Sieverd, como las morfoespecies más frecuentes en este estudio, siendo estas especies de HMA poco frecuentes en muchos de los agroecosistemas del país. Vaast et al. (1997) destacan la respuesta a determinados niveles de P disponible de algunas especies de HMA, en donde se puede presentar una mejor respuesta de una especie a otra, debido a su diferente capacidad para capturar este nutriente del suelo. Por su parte, la relación presentada entre C. pellucida, G. sinuosum, Septoglomus constrictum, Glomus sp. 1 y Acaulospora sp. 1 demostraron una mejor respuesta a la capacidad de captura de [P] en las localidades evaluadas, así como también se destaca la mayor interacción de estas especies con el azufre [S], potasio [K], boro [B], calcio [Ca], cobre [Cu], zinc [Zn], la materia orgánica [MO], conductividad eléctrica [CE] y la capacidad de intercambio catiónico efectiva [CICE], coincidiendo con los reportes que han demostrado que los HMA favorecen significativamente las concentraciones de MO, K, Zn , Ca y Cu, dependiendo de las especies de HMA (Wu et al., 2011).
5. Conclusión
Se reporta por primera vez la ocurrencia de especies de HMA asociadas a cultivos de mango cv. azúcar en las localidades de Santa Marta y Ciénaga, Magdalena, Colombia, encontrándose una alta riqueza de morfoespecies en comparación con otros estudios que se han realizado a nivel mundial. La identificación de morfoespecies claves (dominantes) como Gigasporamargarita y Cetraspora pellucida, encontradas en este estudio, sugieren a estas dos especies como las mejores adaptadas y competitivas en torno a las condiciones en las que se desarrolla el cultivo de mango en las zonas estudiadas. Sin embargo, se necesitan más estudios que podrían ser a nivel molecular, con el fin de identificar la diversidad asociada en las raíces de las plantas, así como su posterior propagación en condiciones de invernadero. Bajo estas condiciones podríamos encontrar nuevos registros de morfoespecies y rescatar toda la riqueza de HMA presente. Esto podría permitir su posterior evaluación con base en la compatibilidad funcional de estos hongos para futuros estudios en diferentes condiciones estresantes.
Contribuciones de los autores
-
Urley Adrian Pérez-Moncada: investigación, metodología, visualización, redacción – borrador original, redacción – revisión y edición.
-
Marlon José Yacomelo-Hernández: conceptualización, investigación, adquisición de fondos, redacción – revisión y edición.
-
Francisco Fabián Carrascal-Pérez: validación, redacción – revisión y edición.
-
Wilmar Alexander Wilches-Ortiz: análisis formal, validación, redacción – revisión y edición.
-
Luciano Ramírez: validación, redacción – revisión y edición.
-
Diana Paola Serralde-Ordoñez: validación, redacción – revisión y edición.
-
Andrea María Peñaranda-Rolon: validación, redacción – revisión y edición.
-
Margarita Ramírez- Gómez: investigación, metodología, conceptualización, adquisición de fondos, redacción – revisión y edición.
Implicaciones éticas
Los autores declaran que no existen implicaciones éticas.
Conflicto de interés
Los autores declaran que no existen conflictos de interés financieros o no financieros que podrían haber influido en el trabajo presentado en este artículo.
Agradecimientos
Agradecemos a la Corporación Colombiana de Investigación Agropecuaria-AGROSAVIA por la financiación de este trabajo, mediante los recursos recibidos en el marco de la ley 1731 de 2014. Los resultados son parte del proyecto “Estrategias de manejo del riego y la nutrición en periodos críticos de sequía para la producción sostenible de mango en zonas productoras de Colombia”.
Referencias
Abdelhalim, T. S., Finckh, M. R., Babiker, A. G., y Oehl, F. (2014). Species composition and diversity of arbuscular mycorrhizal fungi in White Nile state, Central Sudan. Archives of Agronomy and Soil Science, 60(3), 377-391. https://doi.org/10.1080/03650340.2013.793453
Adriano Anaya, M. L., García Ávila, M. A., Ovando Medina, I., De Gyves Córdova, M. G., y Salvador Figueroa, M. (2008). Dinámica de hongos micorrízicos arbusculares en el mango variedad Ataúlfo. Quehacer Científico en Chiapas, 1(6): 25-31. https://www.dgip.unach.mx/images/pdf-REVISTA-QUEHACERCIENTIFICO/QUEHACER-CIENTIFICO-2008-jul-dic/dinamica.pdf
Agronet, (2020). Evaluaciones Agropecuarias Municipales. Grupo de estadística e información sectorial, Oficina de Planeación y Prospectiva, Ministerio de Agricultura y Desarrollo Rural. https://www.agronet.gov.co/Paginas/inicio.aspx
Aguilar Corrales, D. A., Siachoque Bernal, R. F., Otero García, J., Páramo Rocha, G. E., García Márquez, E. R., Escobar Torres, C. H., Porras Rodríguez, P. D., Gómez Blanco, J. A., García Castellanos, L. E., Ayala Pinzón, J. D., Porras Rey, A. M., Olmos Soler, J. R., Enciso Enciso, F. C., y Acosta Latorre, C. P. (2018). Cultivo comercial de mango: identificación de zonas aptas en Colombia, a escala 1:100.000. Unidad de Planificación Rural Agropecuaria [UPRA].
Allen, E. B., Allen, M. F., Egerton-Warburton, L., Corkidi, L., y Gómez-Pompa, A. (2003). Impacts of early and late seral mycorrhizae during restoration in seasonal tropical forest, México. Ecological Applications, 13(6), 1701-1717. https://doi.org/10.1890/02-5309
Belay, Z., Vestberg, M., y Assefa, F. (2014). Mycorrhizal status and AMF community structure of fruit crops from low-input cropping system in Showa Robit, Ethiopia. Ethiopian Journal of Biological Sciences, 13(2), 99-116. https://www.ajol.info/index.php/ejbs/article/view/139802
Boddington, C. L., y Dodd, J. C. (2000). The effect of agricultural practices on the development of indigenous arbuscular mycorrhizal fungi. I. Field studies in an Indonesian ultisol. Plant and Soil, 218/2(1/2), 137-144. https://doi.org/10.1023/A:1014966801446
Brower, J. E., Zar, J. H., y von Ende, C. N. (1990). Field and laboratory methods for general ecology. W.C. Brown Publishers.
Brundrett, M. C., y Tedersoo, L. (2018). Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytologist, 220(4), 1108–1115. https://doi.org/10.1111/nph.14976
Brundrett, M., Melville, L., y Peterson, L. (eds.). (1994). Practical methods in mycorrhiza research. Mycologue Publications.
Chimal-Sánchez, E., Senés-Guerrero, C., Varela, L., Montaño, N. M., García-Sánchez, R., Pacheco, A., Montaño-Arias, S. A., y Camargo-Ricalde, S. L. (2020). Septoglomus mexicanum, a new species of arbuscular mycorrhizal fungi from semiarid regions in Mexico. Mycologia, 112(1), 121-132. https://doi.org/10.1080/00275514.2019.1671147
Cofré, M. N., Soteras, F., del Rosario Iglesias, M., Velázquez, S., Abarca, C., Risio, L., Ontivero, E., Cabello, M. N., Domínguez, L. S., y Lugo, M. A. (2019). Biodiversity of arbuscular mycorrhizal fungi in South America: A review. En M. C. Pagano, y M. A. Lugo (eds.), Mycorrhizal Fungi in South America (pp. 49-72). Springer Cham. https://doi.org/10.1007/978-3-030-15228-4_3
Corazon-Guivin, M. A., Cerna-Mendoza, A., Guerrero-Abad, J. C., Vallejos-Tapullima, A., Carballar-Hernández, S., da Silva, G. A., y Oehl, F. (2019). Nanoglomus plukenetiae, a new fungus from Peru, and a key to small-spored Glomeraceae species, including three new genera in the “Dominikia complex/clades.” Mycological Progress, 18(12), 1395-1409. https://doi.org/10.1007/s11557-019-01522-1
Cortés Mercado, M. M., León Arteaga, R., Ferrera-Cerrato, R., y Cruz Castillo, J. G. (1996). Endomicorrizas vesículo arbusculares del mango (Mangifera indica L.) “Manila” en el estado de Veracruz. Revista Chapingo. Serie horticultura, 2(1), 103-108. https://revistas.chapingo.mx/horticultura/phpscript/download.php?file=completo&id=MzAwMw==
De La Providencia, I. E., de Souza, F. A., Fernández, F., Delmas, N. S., y Declerck, S. (2005). Arbuscular mycorrhizal fungi reveal distinct patterns of anastomosis formation and hyphal healing mechanisms between different phylogenic groups. New Phytologist, 165(1), 261-271. https://doi.org/10.1111/j.1469-8137.2004.01236.x
de Pontes, J. S., Oehl, F., Pereira, C. D., de Toledo Machado, C. T., Coyne, D., da Silva, D. K. A., y Maia, L. C. (2017). Diversity of arbuscular mycorrhizal fungi in the Brazilian’s Cerrado and in soybean under conservation and conventional tillage. Applied Soil Ecology, 117-118, 178-189. https://doi.org/10.1016/j.apsoil.2017.04.023
de Souza, F. A., da Silva, I. R., de Barros Barreto, M. B. B., Oehl, F., Goto, B. T., & Maia, L. C. (2018). Racocetra crispa (Glomeromycotina) delimited by integrative evidence based on morphology, long continuous nuclear rDNA sequencing and phylogeny. Mycological Progress, 17(9), 999-1011. https://doi.org/10.1007/s11557-018-1410-9
FAOSTAT. (2021). Statistics. Food and Agriculture Organization Corporate Statistical Database. http://www.fao.org/statistics/en/
Gerdemann, J. W., y Nicolson, T. H. (1963). Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Transactions of the British Mycological Society, 46(2), 235-244. https://doi.org/10.1016/S0007-1536(63)80079-0
Goto, B. T., y Jobim, K. (2024). Laboratorio de Biología de Micorrizas. https://biologiademicorrizas.wixsite.com/glomeromycota/es
Govindan, M., Rajeshkumar, P. P., Varma, C. K. Y., Anees, M. M., Rashmi, C. R., y Nair, A. B. (2020). Arbuscular mycorrhizal fungi status of mango (Mangifera indica) cultivars grown in typic quartzipsamments soil. Agricultural Research, 9(2), 188-196. https://doi.org/10.1007/s40003-019-00432-8
Jiang, S., Hu, X., Kang, Y., Xie, C., An, X., Dong, C., Xu, Y., y Shen, Q. (2020). Arbuscular mycorrhizal fungal communities in the rhizospheric soil of litchi and mango orchards as affected by geographic distance, soil properties and manure input. Applied Soil Ecology, 152, 103593. https://doi.org/10.1016/j.apsoil.2020.103593
Kindt, R., y Coe, R. (2005). Tree diversity analysis. A manual and software for common statistical methods for ecological and biodiversity studies. World Agroforestry Centre [ICRAF]. https://www.worldagroforestry.org/output/tree-diversity-analysis
Koske, R. E., y Tessier, B. (1983). A convenient permanent slide mounting medium. Mycological Society of America Newsletter, 34(2), 59.
Lekberg, Y., Koide, R. T., Rohr, J. R., Aldrich‐Wolfe, L., y Morton, J. B. (2007). Role of niche restrictions and dispersal in the composition of arbuscular mycorrhizal fungal communities. Journal of Ecology, 95(1), 95-105. https://doi.org/10.1111/j.1365-2745.2006.01193.x
Lin, T.-C., da Silva, G. A., y Oehl, F. (2019). Acaulospora tsugae, a new species in the Glomeromycetes from Taiwan, and a key to species in Acaulosporaceae. Nova Hedwigia, 108(3-4), 475-488. https://doi.org/10.1127/nova_hedwigia/2018/0513
Magurran, A. E. (1988). Ecological diversity and its measurement. Springer Dordrecht. https://doi.org/10.1007/978-94-015-7358-0
Ministerio de Agricultura y Desarrollo Rural [MADR]. (2020). Cadena del Mango. Indicadores e instrumentos Cuarto trimestre. Cifras Sectoriales. Sistema de Información de Gestión y Desempeño de Organizaciones de Cadenas [SIOC]. https://sioc.minagricultura.gov.co/Mango/Documentos/2020-12-31%20Cifras%20Sectoriales.pdf
Mohandas, S. (2012). Arbuscular mycorrhizal fungi benefit mango (Mangifera indica L.) plant growth in the field. Scientia Horticulturae, 143, 43-48. https://doi.org/10.1016/j.scienta.2012.05.030
Mullath, S. K., Błaszkowski, J., Govindan, B. N., Dhaheri, L. al, Symanczik, S., y Al-Yahya’ei, M. N. (2019). Organic farming practices in a desert habitat increased the abundance, richness, and diversity of arbuscular mycorrhizal fungi. Emirates Journal of Food and Agriculture, 31(12), 969-979. https://doi.org/10.9755/ejfa.2019.v31.i12.2057
Oehl, F., de Souza, F. A., y Sieverding, E. (2008). Revision of Scutellospora and description of five new genera and three new families in the arbuscular mycorrhiza-forming Glomeromycetes. Mycotaxon, 106, 311-360.
Oehl, F., Sánchez-Castro, I., da Silva, D. K. A., Santos, V. M., Palenzuela, J., y da Silva, G. A. (2019). Septoglomus nigrum, a new arbuscular mycorrhizal fungus from France, Germany and Switzerland. Nova Hedwigia, 109(1-2), 121-134. https://doi.org/10.1127/nova_hedwigia/2019/0523
Oehl, F., Silva, G. A. da, Goto, B. T., y Sieverding, E. (2011). Glomeromycota: three new genera and glomoid species reorganized. Mycotaxon, 116(1), 75-120. https://doi.org/10.5248/116.75
Oksanen, J., Blanchet, G., Friendly, M., Kindt, R., Legendre, P., Mcglinn, D., Peter, R., Minchin, R. Y., Gavin, L., Simpson, P., Henry, H., Szoecs, E., y Wagner, H. (2020). vegan: Community Ecology Package. R package version 2.5-7. http://CRAN.R-project.org/package=vegan
Öpik, M., Moora, M., Liira, J., y Zobel, M. (2006). Composition of root‐colonizing arbuscular mycorrhizal fungal communities in different ecosystems around the globe. Journal of Ecology, 94(4), 778-790. https://doi.org/10.1111/j.1365-2745.2006.01136.x
Peña-Venegas, C. P., y Vasco-Palacios, A. M. (2019). Endo- and Ectomycorrhizas in Tropical Ecosystems of Colombia. En M. C. Pagano, y M. A. Lugo (eds.), Mycorrhizal Fungi in South America (pp. 111-146). Springer Cham. https://doi.org/10.1007/978-3-030-15228-4_6
Reddy, B., y Bagyaraj, D. J. (1994). Selection of efficient vesicular arbuscular mycorrhizal fungi for inoculating the mango rootstock cultivar ‘Nekkare.’ Scientia Horticulturae, 59(1), 69-73. https://doi.org/10.1016/0304-4238(94)90093-0
Sieverding, E., da Silva, G. A., Berndt, R., y Oehl, F. (2014). Rhizoglomus, a new genus of the Glomeraceae. Mycotaxon, 129(2), 373-386. https://doi.org/10.5248/129.373
Torres-Arias, Y., Furrazola, E., Berbara, R. L. L., Jobim, K., Lima, J. L. R., y Goto, B. T. (2017). Glomus herrerae, a new sporocarpic species of Glomeromycetes from Cuba. Current Research in Environmental & Applied Mycology, 7(3), 155-160. https://doi.org/10.5943/cream/7/3/2
Vaast, P., Zasoski, R. J., y Bledsoe, C. S. (1997). Effects of vesicular-arbuscular mycorrhizal inoculation at different soil P availabilities on growth and nutrient uptake of in vitro propagated coffee (Coffea arabica L.) plants. Mycorrhiza, 6(6), 493-497. https://doi.org/10.1007/s005720050153
van der Heijden, M. G. A., Martin, F. M., Selosse, M., y Sanders, I. R. (2015). Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytologist, 205(4), 1406-1423. https://doi.org/10.1111/nph.13288
Verbruggen, E., Röling, W. F. M., Gamper, H. A., Kowalchuk, G. A., Verhoef, H. A., y van der Heijden, M. G. A. (2010). Positive effects of organic farming on below‐ground mutualists: large‐scale comparison of mycorrhizal fungal communities in agricultural soils. New Phytologist, 186(4), 968-979. https://doi.org/10.1111/j.1469-8137.2010.03230.x
Wu, Q. -S., Li, G. -H., y Zou, Y. N. (2011). Roles of arbuscular mycorrhizal fungi on growth and nutrient acquisition of peach (Prunus persica L. Batsch) seedlings. Journal of Animal and Plant Sciences, 21(4), 746-750. https://www.thejaps.org.pk/Volume/2011/21-4/abstract/16.php
Notas de autor
uperez@agrosavia.co
Información adicional
e-location: e7178